Linkage
Category : NEET
Linkage
Introduction: "When genes are closely present link together in a group and transmitted as a single unit, the is phenomenon is called linkage".
(1) Theories of linkage
(i) Sutton's hypothesis of linkage (1903): The number of groups of genes are equivalent to the number of chromosomes.
(ii) Morgan's hypothesis of linkage (1910): It was given by T. H. Morgan. According to him the genes of homologous parents enter in the same gamete and tend to remain together, which is opposite in heterozygous parents. Linked group are located on the same chromosome and distance between linked group of gene limits the grade of linkage.
(iii) Coupling and repulsion hypothesis: Proposed by Bateson and Punnet (1906) that dominant alleles tend to remain together as well with recessive alleles, called gametic coupling. If dominant and recessive alleles are present in different parents they tend to remain separate and called repulsion. When BBLL and bbll are crossed, the \[{{F}_{1}}\] is BbLl and the test cross of it will show progeny in 7 : 1 : 1 : 7 ratio i.e. BbLl : Bbll : bbLl : bbll (coupling) when BBll is crossed with bbLL the \[{{F}_{1}}\] is BbLl or the test cross progeny will show 1 : 7 : 7 : 1 ratio i.e., BbLl : Bbll : bbLl : bbll (repulsion). Coupled and repulsed genes are known as linked genes. Linkage has coupling phase and repulsion phase. In coupling phase both the linked genes have their dominant alleles in one chromosome and recessive alleles in other chromosomes. The heterozygotes with such constitution is called cis heterozygote. Cis-arrangement is a original arrangement. Which form two types of gametes as (AB) and (ab). In Human X–chromosomes carry 102 genes and Y
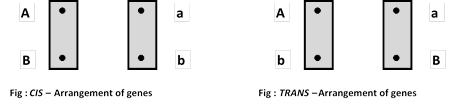
In repulsion phase the normal alleles as well as mutant alleles lie in opposite chromosomes of the homologous pair, such heterozygote is called as trans heterozygote. It is not original arrangement, caused due to crossing over, which form two types of gametes as (Ab) and (aB).
(iv) Chromosomal hypothesis of linkage: It was given by Morgan and Castle. According to them linked genes are bound by chromosomal material and are transmitted as a whole.
(2) Types of linkage: Depending upon the absence or presence of nonparental or new combination of linked genes, linkage has been found to be complete or incomplete.
(i) Complete linkage: Such cases in which linked genes are transmitted together to the offsprings only in their original or parental combination for two or more or several generations exhibit complete linkage. In such cases the linked genes do not separate to form the new or non-parental combinations. This phenomenon is very rare. Some characteristics in males of Drosophila are found to exhibit complete linkage.
(ii) Incomplete linkage: In majority of cases, the homologous chromosomes undergo breakage and reunion during gametogenesis. During reunion the broken pieces of the chromatids are exchanged, producing some nonparental or new combinations. Therefore, the linkage is rendered incomplete. The phenomenon of interchange of chromosome segments between two homologous chromosomes is called crossing over. Incomplete linkage is very common and has been studied in almost all the organisms.
(3) Linkage groups: All the genes which are linked with one another, form a linkage group. Since linked genes are present in the same chromosome, the number of linkage group in an animal or plant is equal to the haploid number of chromosomes present in its cells. This hypothesis was given by Sutton and was proved by experiments on Drosophila by T.H. Morgan.
|
Examples |
Linkage groups |
|
Drosophila |
There are four linkage groups corresponding to the four pairs of chromosomes |
|
Zea mays |
The ten chromosome pairs of maize correspond with its ten linkage groups |
|
Pisum sativum |
The garden pea plant has seven pairs of chromosome and the same number of linkage groups |
|
Man |
Man has 23 linkage groups corresponding to 23 pairs of chromosomes |
(4) Strength of linkage: The strength of linkage between any two pairs of linked genes of a chromosome depend upon the distance between them. Closely located genes show strong linkage, while genes widely located show weak linkages.
(5) Factor affected to linkage: Linkage is affected by the following factors.
(i) Distance: Closely located genes show strong linkage while genes widely located show weak linkage.
(ii) Age: With increasing age the strength of linkage decreases.
(iii) Temperature: Increasing temperature decreases the strength of linkage.
(iv) X-rays: X-rays treatment reduces the strength of linkage.
(6) Significance of linkage
(i) Due to linkage new recombinants are formed.
(ii) It helps in maintaining the valuable traits of a newly developed variety.
(iii) It helps locating genes on chromosome.
(iv) It disallows the breeders to combine all the desirable traits in a single variety.
Important Tips
Crossing Over
Introduction: The process by which exchange of chromosomal segment take place is called crossing over. Crossing over may be defined as "the recombination of linked genes" brought about as a result of interchange of corresponding parts between the chromatid of a homologous pair of chromosomes, so as to produce new combination of old genes. The term was given by Morgan and Cattle. Janssen (1909) observed chiasmata during meiosis-I (Prophase). Morgan proposed that chiasmata lead to crossing over by breakage and reunion of homologous chromosomes. Crossing over results in new combination while non-cross over result in parental type, which leads to variations.
(1) Kinds of crossing over
(i) Somatic crossing over: It is found in somatic cells e.g., Curt stern in Drosophila and Potnecorvo in Aspergillus nidulans shown somatic crossing over i.e. mitotic crossing over.
(ii) Germinal crossing over: It is found in germinal cells during gametogenesis. This is also known as meiotic crossing over.
(iii) Single cross over: It takes place at one point only on the non-sister chromatids, only 2 chromatids are involved.
(iv) Double cross over: It is the formation of 2 chiasmata in the same chromosome independent of each other. In a double cross over the genes lying outside the crossed regions will retain their original association.
(v) Multiple cross over: It is formed when more than 2 chiasmata are formed. It is very rare.
(vi) Two-stranded crossing over: It takes place before splitting of homologous chromosomes so all the four resultants are recombinants.
(vii) Four stranded crossing over: It takes place after splitting of homologous chromosomes only 2 non-sister chromatids take part in crossing over resulting in 2 parental and 2 recombinant types.
(2) Crossing over and chiasma: There are two views extended to explain the relationship between crossing over and chiasma formation. They are summarised here under.
(i) Chiasma type theory: According to Janssen, 1909 the act of crossing over is followed by chiasma formation. He suggests that the crossing over takes place at the pachytene stage and the chiasma appear at diplotene.
(ii) Classical theory: According to Sharp, 1934, crossing over is the result of chiasma formation. According to this view, the chiasma are organised at pachytene and crossing over takes place at diplotene stage. On the basis of evidence available from molecular biology, that is untenable and hence rejected.
(3) Mechanism of crossing over: There are different views put forward to explain the mechanism of crossing over.
(i) Copy choice hypothesis: According to Belling, 1928 the chromomeres represent the genes joined by interchromomeric regions. The chromomeres duplicate first and then the interchromomeric regions. The synthesis of these regions may occur in such a way that the chromomeres of the chromatid of a homologue get connected of the chromatid of the other homologue at a specific location. As a result, the adjacent chromatids of a pair of homologue are exchanged.
(ii) Precocity hypothesis: According to Darlington, the pairing of homologues occurs to avoid singleness of a chromosome. The pairing need of a chromosome could be nothing less than the replication of DNA. The crossing over takes place due to torsion on chromosome created by coiling of the two homologues around each other.
In fact, the crossing over is the event which, precisely at molecular level, results in the formation of a hybrid DNA molecule. Such models have been proposed by White house, 1963 as also by Holliday, 1964. These models mainly elaborate the mechanism of breakage and reunion of DNA helicase.
(4) Cross over value: The percentage of crossing over varies in different materials. The frequency of crossing over is dependent upon the distance of two genes present on a chromatid.
(5) Coincidence: Coincidence or coefficient of coincidence is inverse measure of interface and is expressed as the ratio between the actual number of double cross over and the expected number of such double cross. That is:
\[\text{Coincidence}=\,\frac{\text{Actual number of double cross over}}{\text{Expected number of double cross over}}\]
(6) Factors controlling frequency of crossing over: Primarily, frequency of crossing over is dependent upon the distance between the linked genes, but a number of genetic, environmental and physiological factors also affect it. These are:
(i) Temperature: High and low temperature increase the frequency of crossing over.
(ii) X-ray: Muller has discovered that exposure to X-ray and other radiations increases the frequency of crossing over.
(iii) Age: The frequency of crossing over decreases with increasing age in female Drosophila.
(iv) Chemicals: Certain chemicals which act as mutagens do affect the frequency of crossing over. Gene mutations may affect the frequency of crossing over. Some increase the frequency, whereas some may decrease it.
(v) Sex: Crossing over in Drosophila males is negligible. Males of mammals also exhibit little crossing over. In silk-moth, crossing over does not occur in females.
(vi) Chiasmata formation: Chiasmata formation at one point discourages chiasmata formation and crossing over in the vicinity. This phenomenon is known as interference.
(vii) Inversions: Inversions of chromosome segments suppresses crossing over.
(viii) Distance: Distance between the linked genes is the major factor which controls the frequency of crossing over. The chances of crossing over between distantly placed genes are much more than between the genes located in close proximity.
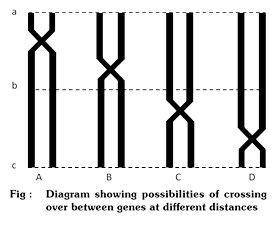
Figure depicts that chance of crossing over between a and c are double as compared to the chances between a and b or b and c.
(ix) Cytoplasm: Factor for crossing over is present in cytoplasm and is inherited to the offspring.
(x) Nutritional effect: Crossing over frequencies are affected by concentration of metallic ions, such as calcium and magnesium.
(xi) Genotypic effect: Crossing over frequencies between the same two loci in different strains of the same species show variation because of numerous gene differences.
(xii) Chromosome structure effect: Changes in the order of genes on a chromosome produced by chromosomal aberrations usually act as cross over suppressors.
(xiii) Centromere effect: Genes present close to the centromere region show reduced crossing over.
(xiv) Interference: If there are two doubles crossovers, then one crossover tries to influence the other by suppressing it. This phenomenon is called as interference. Due to this phenomenon, the frequency of crossing over is always lower than the expected.
(7) Significance of crossing over: This phenomenon is of great biological significance, which are as under:
(i) It gives evidence that the genes are linearly arranged on a chromosome. Thus, it throws light on the nature and working of the genes.
(ii) It provides an operational definition to a gene. It is deemed as the smallest heritable segment of a chromosome in the interior of which no crossing over takes place.
(iii) The crossing over is helpful in the chromosomal mapping. The percentage of crossing over is proportional to the distance between two genes.
(iv) It is the main cause of genetic variations. It's occurrence during the act of meiosis produces variations in the heritable characters of the gametes.
(v) This phenomenon has also found it's utility in breeding and evolving new varieties. The linkage of undesirable characters can be broken by temperature treatment, using X-ray or chemicals. Thus, new recombinants can be prepared.
Important Tips




You need to login to perform this action.
You will be redirected in
3 sec

