Krebs Cycle and ETS
Category : NEET
Krebs Cycle and ETS
(1) Oxidative decarboxylation / Formation of acetyl CoA.
(2) Kreb's cycle / TCA cycle / Citric acid cycle.
(3) Electron Transport System
(1) Oxidative decarboxylation of pyruvic acid : If sufficient O2 is available, each 3-carbon pyruvate molecule \[\left( C{{H}_{3}}COCOOH \right)\] enters the mitochondrial matrix where its oxidation is completed by aerobic means. It is called gateway step or link reaction between glycolysis and Kreb's cycle. The pyruvate molecule gives off a molecule of CO2 and releases a pair of hydrogen atoms from its carboxyl group (-COOH), leaving the 2 carbon acetyl group \[(C{{H}_{3}}CO)\]. The reaction is called oxidative decarboxylation, and is catalyzed by the enzyme pyruvate dehydrogenase complex (decarboxylase, TPP, lipolic acid, transacetylase, Mg2+) . During this reaction, the acetyl group combines with the coenzyme A (CoA) to form acetyl coenzyme A with a high energy bond\[\left( C{{H}_{3}}CO-CoA \right)\]. Most of the free energy released by the oxidation of pyruvate is captured as chemical energy in high energy bond of acetyl coenzyme A. From a pair of hydrogen atoms released in the reaction, to electrons and one H+ pass to\[NA{{D}^{+}}\], forming, \[NAD{{H}^{+}}{{H}^{+}}\]. The NADH forms 3 ATP molecules by transferring its electron over ETS described ahead.
Decarboxylation and dehydration :
**TPP=Thiamine pyrophosphate
**LAA=Lipoic acid amide
Acetyl CoA is a common intermediate of carbohydrate and fat metabolism. Latter this acetyl CoA from both the sources enters Kreb's cycle. This reaction is not a part of Kreb's cycle.
(2) Kreb's cycle / TCA cycle / Citric acid cycle
(i) Discovery : This cycle has been named after the German biochemist in England Sir Hans Krebs who discovered it in 1937. He won Noble Prize for this work in 1953. Krebs cycle is also called the citric acid cycle after one of the participating compounds.
(ii) Site of occurrence : It takes place in the mitochondrial matrix.
(iii) Kreb’s cycle
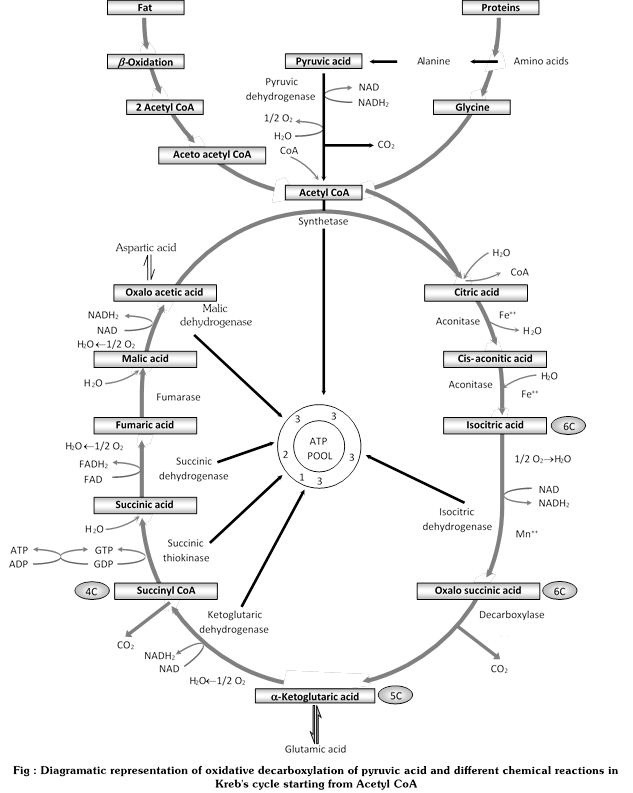
(iv) Enzymes of Kreb's cycle
|
Step |
Enzyme |
(Location in mitochondria) |
Coenzyme(s) and cofactor (s) |
Inhibitor(s) |
Type of reaction catalyzed |
|
(a) |
Citrate synthetase |
Matrix space |
CoA |
Monofluoro-acetyl- CoA |
Condensation |
|
(b) |
Aconitase |
Inner membrane |
\[F{{e}^{2+}}\] |
Fluoroacetate |
Isomerization |
|
(c) |
Isocitrate dehydrogenase |
Matrix space |
\[NA{{D}^{+}},\text{ }NAD{{P}^{+}},\text{ }M{{g}^{2+}},\text{ }M{{n}^{2+}}\] |
ATP |
Oxidative decarboxylation |
|
(d) |
\[\alpha \]-ketoglutarate dehydrogenase complex |
Matrix space |
TPP,LA,FAD,CoA, NAD+ |
Arsenite,Succinyl-CoA, NADH |
Oxidative decarboxylation |
|
(e) |
Succinyl-CoA synthetase |
Matrix space |
CoA |
- |
Substrate level phosphorylation |
|
(f) |
Succinate dehydrogenase |
Inner membrane |
FAD |
Melonate, Oxaloacetate |
Oxidation |
|
(g) |
Fumarase |
Matrix space |
None |
- |
Hydration |
|
(h) |
Malate dehydrogenase |
Matrix space |
NAD+ |
NADH |
Oxidation |
(v) Steps in Kreb's cycle : Kreb's cycle consists of 8 cyclic steps, producing an equal number of organic acids. Each step is catalyzed by a specific enzyme. In Kreb's cycle, the entrant molecule is 2-carbon acetyl CoA and the receptor molecule is 4- carbon oxaloacetate.
(a) condensation : Acetyl coenzyme A reacts in the presence of water with the oxaloacetate normally present in a cell, forming 6-carbon citrate and freeing coenzyme A for reuse in pyruvate oxidation. The high-energy bond of acetyl CoA provides the energy for this reaction. The reaction is catalyzed by the citrate synthetase enzyme. The citrate has 3-carboxyl group. Hence, Krebs cycle is also called tricarboxylic acid cycle, or TCA cycle after its first product.
\[\text{Oxaloacetate}+\text{Acetyl}\,\text{CoA }+{{\text{H}}_{\text{2}}}O\underset{\text{Synthetase}}{\mathop{\xrightarrow{\text{Citrate}}}}\,\text{Citrate}+CoA\,(\text{Reused})\]
(b) Reorganisation (Dehydration) : Citrate undergoes reorganisation in the presence of an enzyme, aconitase , forming 6-carbon cisaconitate and releasing water.
\[\text{Citrate}\xrightarrow{\text{Aconitase}}\text{Cisaconitate}+{{H}_{2}}O\]
(c) Reorganisation (Hydration) : Cisaconitate is further reorganised into 6-carbon isocitrate by the enzyme, aconitase, with the addition of water.
\[\text{Cisaconitate}+{{H}_{2}}O\xrightarrow{\text{Aconitase}}\text{Isocitrate}\]
(d) Oxidative decarboxylation I : This is a two stage process :
Stage I : Hydrogen atoms from isocitric acid react with NAD to form NAD. 2H forming oxalosuccinic acid. The pair of hydrogen atoms give two electrons and one H+ to NAD+ forming NADH +H+. The enzyme isocitrate dehydrogenase catalyses the reaction in the presence of Mn2+. NADH generates ATP by transferring its electron over the ETS.
\[\text{Isocitric}\,\text{acid}+NAD\underset{M{{n}^{2+}}}{\mathop{\xrightarrow{\text{Isocitric}\,\text{dehydrogenase}}}}\,\text{Oxalosuccinic}\,\text{acid}+NAD.2H(or\,NADPH.2H)\]
Stage II : Decarboxylation of oxalosuccinic acid occurs forming \[\alpha \]-ketoglutaric acid, which is a first 5-C carbon molecule of Kreb's cycle.
\[\text{Oxalosuccinic}\,\text{acid}\underset{{}}{\mathop{\underset{M{{n}^{2+}}}{\mathop{\xrightarrow{\text{Carboxylase}}}}\,\alpha -\text{Ketoglutaric}\,\text{acid}+C{{O}_{2}}}}\,\]
(e) Oxidative decarboxylation II : This is also a 2 stage process :
Stage I : Coenzyme A reacts with \[\alpha \]-ketoglutarate, forming 4-carbon succinyl-coenzyme A and releasing CO2 and a pair of hydrogen atoms. The reaction is catalysed by \[\alpha \]-ketoglutarate dehydrogenase complex enzyme. the pair of hydrogen atoms pass two electrons and one H+ to NAD+, forming NADH + H+
\[\alpha -\text{ketoglutarate}+CoA+NA{{D}^{+}}\underset{\text{dehydrogenase}}{\mathop{\xrightarrow{\alpha -\text{ketoglutarate}}}}\,\text{Succinyl}-CoA+C{{O}_{2}}+NADH+{{H}^{+}}\]
Stage II : Succinyl -coenzyme A splits into 4-carbon succinate and coenzyme A with the addition of water. The coenzyme A transfers its high energy to a phosphate group that joins GDP (Guanosine diphosphate), forming GTP (Guanosine triphosphate). The latter is an energy carrier like ATP. This is the only high-energy phosphate produced in the Krebs cycle. The stage 2 reaction is catalysed by succinyl-CoA synthetase enzyme. The formation of GTP is called substrate level phosphorylation.\[\]
\[\text{Succinyl}-CoA+{{H}_{2}}O+GDP/ADP\underset{\text{Synthetase}}{\mathop{\xrightarrow{\text{Succinyl}-CoA}}}\,\text{Succinate}+CoA+GTP/ATP\]
In a plant cell, this reaction produce ATP from ADP and GTP from GDP or ITP (Inosine triphosphate) in animals.
Oxygen to oxidize a carbon atom to CO2 is taken in steps 4 and 5 from a water molecule.
(f) Dehydrogenation : This process converts succinate into 4-carbon fumarate with the aid of an enzyme, succinate dehydrogenase, and liberates a pair of hydrogen atoms. The latter pass to FAD+ (Flavin adenine dinucleotide), forming FADH2. Hydrogen is carried by FAD in the form of whole atoms.
Succinate+FAD+![]() Fumarate +FADH2
Fumarate +FADH2
(g) Hydration : This process changes fumarate into 4-carbon maltate in the presence of water and an enzyme, fumarase.
Fumarate +H2O ![]() Maltate
Maltate
(h) Dehydrogenation : This process restores oxaloacetate by removing a pair of hydrogen atoms from maltate with the help of an enzyme maltate dehydrogenase. The pair of hydrogen atoms pass two electrons and one H+ to NAD+ , forming NADH+H+.
Maltate +NAD+![]() Oxaloacetate \[+NADH\text{ }+{{H}^{+}}\]
Oxaloacetate \[+NADH\text{ }+{{H}^{+}}\]
Oxaloacetate combines with acetyl coenzyme A to form citrate, and so the cycle continues.
(vi) Summary of Kreb's cycle
(a) All the enzymes, reactants, intermediates and products of TCA cycle also are found in aqueous solution in the matrix, except the enzyme a-ketoglutarate dehydrogenase and succinate dehydrogenase which are located in the inner mitochondrial membrane. Both are called mitochondrial marker enzyme.
(b) Oxidation of one mole of acetyl CoA uses 4 molecules of water and releases one molecule of water.
(c) Liberates 2 molecules of carbon dioxide.
(d) Gives off 4 pairs of hydrogen atoms.
(e) Produces one GTP/ ATP molecule during the formation of succinate.
(f) One mole of acetyl CoA gives 12 ATP during oxidation in Krebs cycle.
(g) Regenerates oxaloacetate used in last cycle for reuse.
The above summary is for one molecule of acetyl coenzyme A. There are two acetyl coenzyme A molecules formed from one molecule of glucose by glycolysis and oxidative decarboxylation of pyruvate. The entire Krebs cycle may be represented by the following equation -
\[2\text{Acetyl}\,\text{coenzyme}\,A+8{{H}_{2}}O+6NA{{D}^{+}}+2FA{{D}^{+}}+2GDP/ADP+2Pi\] \[\to 4C{{O}_{2}}+2{{H}_{2}}O+6\,NADH+2FAD{{H}_{2}}+2GTP/ATP+6{{H}^{+}}\]
(vii) Difference between Glycolysis and Kreb's cycle
| Glycolysis | Kreb?s cycle |
|
It takes place in the cytoplasm. |
It takes place in the matrix of mitochondria. |
|
It occurs in aerobic as well as anaerobic respiration. |
It occurs in aerobic respiration only. |
|
It consists of 9 steps. |
It consists of 8 steps. |
|
It is a linear pathway. |
It is a cyclic pathway. |
|
It oxidised glucose partly, producing pyruvate. |
It oxidises acetyl coenzyme A fully. |
|
It consumes 2 ATP molecules. |
It does not consume ATP |
|
It generates 2 ATP molecules net from 1 glucose molecules. |
It generates 2 GTP/ATP molecules from 2 succinyl coenzyme A molecules. |
|
It yields 2 NADH per glucose molecule. |
It yields 6 NADH molecules and 2 FADH2 molecules from 2 acetyl coenzyme A molecules. |
|
It does not produce CO2. |
It produces CO2. |
|
All enzyme catalysing glycolytic reactions are dissolved in cytosol. |
Two enzymes of Krebs cycle reactions are located in the inner mitochondrial membrane, all others are dissolved in matrix. |
(viii) Product form during aerobic respiration by Glycolysis and Kreb’s cycle.
(a) Total formation of ATP
| ATP formation in Glycolysis | |||||
|
|
Steps |
Product of reactions |
In terms of ATP |
||
|
ATP formation by substrate phosphorylation |
1, 3-diphosphoglyceric acid (2 moles) ® 3 phosphoglyceric acid (2 moles) Phosphoenolpyruvic acid (2 moles) ® Pyruvic acid (2 moles) |
2 ATP 2 ATP |
2 ATP 2 ATP |
||
|
|
|
Total |
4 ATP |
||
|
ATP formation by oxidative phosphorylation or ETC |
1, 3 - disphosphoglyceraldehyde (2 moles) 1, 3 - diphosphoglyceric acid (2 moles) |
2 NADH2 |
6 ATP |
||
|
|
Total ATP formed |
4 + 6 ATP = |
10 ATP |
||
|
ATP consumed in Glycolysis |
Glucose (1 mole) ® Glucose 6 phosphate (1 mole) Fructose 6 phosphate (1 mole) ® Fructose 1, 6-diphosphate (1 mole) |
- 1 ATP - 1 ATP |
- 1 ATP
- 1 ATP |
||
|
|
|
Total |
2 ATP |
||
|
|
Net gain of ATP = total ATP formed - Total ATP consumed |
10 ATP - 2ATP |
8 ATP |
||
| ATP formation in Kreb?s cycle | |||||
|
ATP formation by substrate phosphorylation |
Succinyl CoA (2 mols) ® Succinic acid (2 mols) |
2 GTP |
2 ATP |
||
|
|
|
Total |
2 ATP |
||
|
ATP formation by oxidative phosphorylation or ETC |
Pyruvic acid (2 mols) ® Acetyl CoA (2 mols) Isocitric acid (2 mols) ® Oxalosuccinic acid (2 mols) a-Ketoglutaric acid (2 mols) ® Succinyl CoA (2 mols) Succinic acid (2 mols) ® Fumaric acid (2 mols) Malic acid (2 mols) ® Oxaloacetic acid (2 mols) |
2 NADH2
2 NADH2
2 NADH2
2 FADH2
2 NADH2 |
6 ATP
6 ATP
6 ATP
4 ATP
6 ATP |
||
|
|
|
Total |
28 ATP |
||
|
|
Net gain in Kreb?s cycle (substrate phosphorylation + oxidative phosphorylation) |
2ATP + 28 ATP |
30 ATP |
||
|
Net gain of ATP in glycolysis and Kreb?s cycle |
Net gain of ATP in glycolysis + Net gain of ATP in Kreb?s cycle |
8 ATP + 30 ATP |
38 ATP |
||
|
Over all ATP production by oxidative phosphorylation or ETC |
ATP formed by oxidative phosphorylation in glycolysis + ATP formed by oxidative phosphorylation or ETC. |
6 ATP + 28 ATP |
34 ATP |
||
22 ATP produced by oxidation of NADH2 and FADH2 in Kreb’s cycle and 6 ATP comes from oxidative decarboxylation of pyruvic acid.
(b) Formation and use of water
| Formation of water molecules | ||
|
Formation of water molecules in glycolysis |
2 phosphoglyceric acid (2 mols) \[\xrightarrow{-{{H}_{2}}O}\] 2 phosphoenol pyruvic acid (2 mols) 1, 3-diphosphoglyceraldehyde \[\xrightarrow{-{{H}_{2}}O}\] 1, 3 diphosphoglyceric acid |
2H2O
2H2O |
|
|
Total water molecules formed in glycolysis |
4H2O |
|
Formation of water molecules in kreb?s cycle |
One molecule of water in each of the five oxidation reactions (these reactions occur twice as there are two molecules of pyruvic acid). Other than oxidation reaction Citric acid (2 mols) ® Cis-aconitic acid (2 mols) |
10 H2O
2H2O |
|
|
Total water molecules formed in Kreb?s cycle |
12 H2O |
|
|
Total water molecules formed in aerobic respiration (Glycolysis + Kreb?s cycle) |
16 H2O |
| Use of water molecules | ||
|
Use of water in Glycolysis |
3-phosphoglyceraldehyde (2 mols) \[\xrightarrow{+{{H}_{2}}O}\] 1, 3 diphosphoglyceric acid (2 mols) |
2H2O |
|
|
Total water molecule used in glycolysis |
2H2O |
|
Use of water in Kreb?s cycle |
Oxaloacetic acid (2 mols) \[\xrightarrow{+{{H}_{2}}O}\] Citric acid (2 mols) Cis aconitic acid (2 mols) \[\xrightarrow{+{{H}_{2}}O}\] Isocitric acid (2 mols) Succinyl CoA (2 mols) \[\xrightarrow{+{{H}_{2}}O}\] Succinic acid (2 mols) Fumaric acid (2 mols) \[\xrightarrow{+{{H}_{2}}O}\] Malic acid (2 mols) |
2H2O 2H2O 2H2O 2H2O |
|
|
Total water molecules used is Kreb?s cycle |
8H2O |
|
|
Total water molecules used in aerobic respiration (Glycolysis + Kreb?s cycle) |
10H2O |
|
Net gain of water molecules in aerobic respiration |
Number of water molecules formed - Number of water molecules used = ( 16 H2O - 10H2O) |
6H2O |
(c) Evolution of carbon dioxide
|
Pyruvic acid (2 mols) \[\xrightarrow{-C{{O}_{2}}}\] Acetyl CoA (2 mols) Oxalosuccinic acid \[\xrightarrow{-C{{O}_{2}}}\] a ketoglutaric acid (2 mols) a Ketoglutaric acid (2 mols) \[\xrightarrow{-C{{O}_{2}}}\] Succinyl CoA (2 mols) |
2CO2 2CO2 2CO2 |
|
Total CO2 molecules released in aerobic respiration |
6CO2 |
(d) Use of O2 (Oxygen)
|
Use of oxygen in Glycolysis |
1, 3-diphosphoglyceraldehyde (2mols) \[\xrightarrow{+\frac{1}{2}{{O}_{2}}}\] 1, 3-diphosphoglyceric acid (2 mols) |
1O2 |
|
Use of oxygen in Kreb?s cycle |
Five oxidation reactions of Kreb?s cycle (2 times) |
5O2 |
|
|
Total O2 molecules required for aerobic respiration |
6O2 |
(ix) Energy storage and energy transfer : In respiration energy released takes in the form of chemical energy, stored in a form called ATP . Energy transfer of biological oxidation hinges on the formation of labile high energy phosphate bonds of ATP. Nicotinamide adenine dinucleotide phosphate (NAD), Flavin adenine dinucleotide (FAD), Guanosine triphosphate are also the product of respiration and converted to ATP by electron transport system.
(a) Adenosine triphosphate : An energy intermediate :
There are several compounds like NAD, FAD, GTP and ATP are known as energy yielding compounds. The best known, and probably the most important of these are adenosine triphosphate (ATP). It serves as the energy currency of the cells.
Structure of ATP : Adenosine triphosphate is a nucleotide consisting of three main constituents;
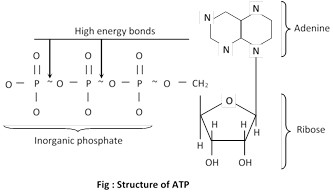
(a) A nitrogen contain purine base
(b) A five carbon sugar ribose
(c) Three inorganic phosphate groups
The bonds attaching the last two phosphate to the rest of the molecule are high energy bonds (~) contain more than twice the energy of an average chemical bond.
ADP further hydrolysed to AMP and inorganic phosphate, releasing 7.3 Kcal energy per molecule (of ATP). Above process represented by following reactions.
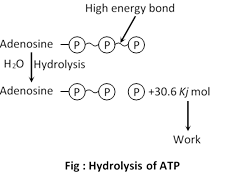
\[\text{Adenosine}\,\text{triphosphate}\xrightarrow{\text{hydrolysis}}\text{Adenosine}\,\text{diphosphate}\,(ADP)\underset{\text{hydrolysis}}{\mathop{+Pi+}}\,7.3Kcal.....\]
\[\text{Adenosine}\,\text{diphosphate}\to \text{Adenosine}\,\text{monophosphate(AMP)}+Pi+7.3Kcal.\]
Phosphorylation : The ATP hydrolysis reactions are reversible because ATP are synthesized from ADP, Pi and energy (take up for the bond formation). The addition of phosphate group to ADP and AMP called phosphorylation. Energy required for the bond formation is equal to the energy released in hydrolysis.
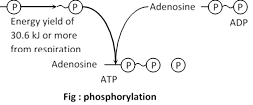
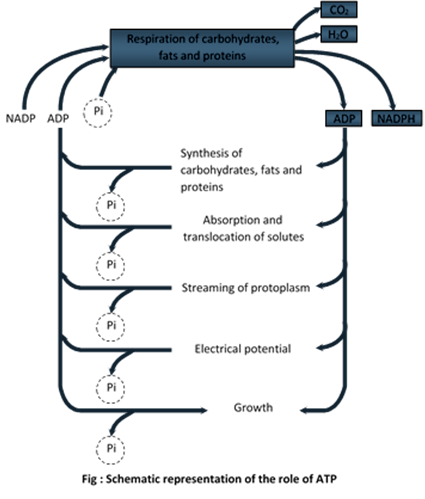
The significant role of ATP as an intermediate energy transfer compound
(a) Synthesis of carbohydrates, proteins, fats, etc.
(b) Translocation of organic food.
(c) Absorption of organic and inorganic food.
(d) Protoplasmic streaming.
(e) Growth.
(b) Nicotinamide dinucleotide phosphate/ Nicotinamide dinucleotide (NADP/NAD) : It is called universal hydrogen acceptor, produced during aerobic respiration (glycolysis+ Kreb's cycle) and also in anaerobic respiration, work as coenzyme in ATP generation Via electron transport system. NADP have one additional phosphate.
Structure of NAD = Nicotinamide-adenine-dinucleotide (formerly called coenzyme I or CO-I Diphosphopyridine dinucleotide = DPN) is shown below:
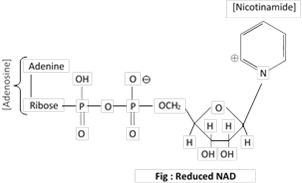
It plays a crucial role in dehydrogenation processes. Some dehydrogenases do not work with NAD, but react with NADP (Nicotinamide adenine dinucleotide phosphate). Formerly called Coenzyme II or Triphosphopyridine nucleotide = TPN Nicotinamide is a vitamin of B group.
First NAD and NADP both functions as hydrogen acceptors. Later H ions and electrons (e_) from these are transported through a chain of carriers and after being released at the end of a chain react with O2 and from H2O (see Electron Transport chain). During the release of 2 electron from 2H+ atoms from NAD. 2H and their reaction with O2 to form water, 3 ATP molecules are synthesized.
Important Tips
(i) NADH2 may be used in some other metabolic pathway.
(ii) NADH2 do not always produces 3 ATP molecules. More active muscle cells produces more while less active fat cell produce less ATP molecules.
(iii) Not all the proteins are routed through F0 -F1 channel.
(iv) So realistic aerobic respiratory efficiency ranges between 22% to 38%, as realistic power limit is 21 ATP molecule.
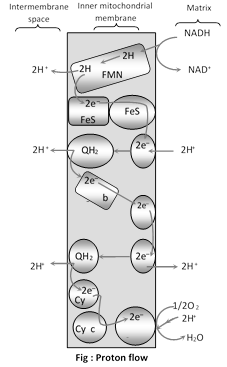
(3) Electron transport system : The electron transmitter system is also called electron transport chain (ETC), or cytochrome system (CS), as four out of these seven carriers are cytochrome. It is the major source of cells energy, in the respiratory breakdown of simple carbohydrates intermediates like phosphoglyceraldehyde, pyruvic acid, isocitric acid, \[\alpha -\]ketoglutaric acid, succinic acid and malic acid are oxidised. The oxidation in all these brought about by the removal of a pair of hydrogen atoms (2H) from each of them. This final stage of respiration is carried out in ETS, located in the inner membrane of mitochondria (in prokaryotes the ETS is located in mesosomes of plasma membrane). The system consists of series of precisely arranged seven electron carriers (coenzyme) in the inner membrane of the mitochondrion, including the folds or cristae of this membrane. These seven electron-carriers function in a specific sequence and are:
Nicotinamide adenine dinucleotide (NAD), Flavin mononucleotide (FMN), Flavin adenine dinucleotide (FAD), Co-enzyme-Q or ubiquinone, Cytochrome-b, Cytochrome-c, Cytochrome-a and Cytochrome-a3,

The first carrier in the chain is a flavoprotein which is reduced by NADH2. Coenzyme passes these electron to the cytochromes arranged in the sequence of b-c-a-a3, finally pass the electron to molecular oxygen. In this transport, the electrons tend to flow from electro-negative to electro-positive system, so there is a decrease in free energy and some energy is released so amount of energy with the electrons goes on decreasing. During electron-transfer, the electron-donor gets oxidised, while electron-acceptor gets reduced so these transfers involve redox-reaction and are catalysed by enzymes, called reductases. Oxidation and reduction are complimentary. This oxidation-reductiion reaction over the ETC is called biological oxidation.
\[\text{Electron}-\text{donor}\to {{\text{e}}^{\text{-}}}+\text{electron}-\text{acceptor}\]
here, electron-donor and electron -acceptor form redox pair.
During the electron transfers, the energy released at some steps is so high that ATP is formed by the phosphorylation of ADP in the presence of enzyme ATP synthetase present in the head of F1-particles present on the mitochondrial crista. This process of ATP synthesis during oxidation of coenzyme is called oxidative phosphorylation, so ETS is also called oxidative phosphorylation pathways.
\[\text{ADP}+\text{Pi}\xrightarrow{\text{ATP Synthetase}}\text{ATP}\]
From the cytochrome a3, two electrons are received by oxygen atom which also receives two proton (H+) from the mitochondrial matrix to form water molecule. So the final acceptor electrons is oxygen. So the reaction
\[{{H}_{2}}+\frac{1}{2}{{O}_{2}}\to {{H}_{2}}O\](called metabolic water) is made to occur in many steps through ETC, so the most of the energy can be derived into a storage and usable form.
(i) Two route systems of ETC: The pairs of hydrogen atoms from respiratory intermediates are received either by NAD+ or FAD coenzymes which becomes reduced to NADH2 and FADH2. These reduced coenzyme pass the electrons on to ETC. Thus, regeneration of NAD+ or FAD takes place in ETC. There are two routes ETC :
(a) Route 1 : NADH2 passes their electrons to Co-Q through FAD . In route 1 FAD is the first electron carrier. 3 ATP molecules are produced during the transfer of electron on following steps :
NAD to FAD
Cyt b to Cyt c and
Cyt a to Cyt a3
(b) Route 2 : FADH2 passes their electron directly to FAD. 2 ATP molecules are produced during the transfer of electron on following steps.
Cyt b to Cyt c and
Cyt a to Cyt a3
(ii) Structure of mitochondria in relation to oxidative function : On inner side of mitochondria elementary particles or F0-F1 complex of ATPase complex or elementary particle (oxysomes) are found. Previously it was considered that elementary particles contain all the enzyme of oxidative phosphorylation and electron transport chain.
Component of electron transport chain are located in the inner membrane in the form of respiratory chain complexes. For complexes following theories are given :
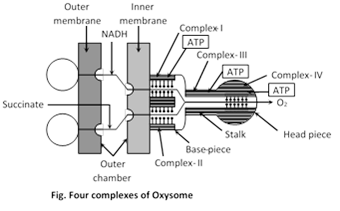
(a) Four complex theory : According to Devid green electron transport chain contains 4 complexes-
Complex I : Comprises NADH dehydrogenase and its 6 Iron Sulphur centers (Fe-S).
Complex II : Consists of Succinate dehydrogenase and its 3 Iron Sulphur centers.
Complex III : Consists of cytochrome b and c, and a specific Iron-Sulphur centers.
Complex IV : Comprises cytochromes a and a3.
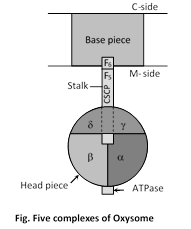
(b) Five complex theory : According to Hatefi, (1976), Complex I to Complex IV are related to the electron transport.
Complex II : Succinate : CoQ reductase
Complex III : Reduced CoQ (CoQH2) : cytochrome C reductase
Complex IV : Cytochrome C oxidase
Complex V : ATPase
Cytochrome C and Q are mobile components of the respiratory chain.
(iii) Oxidative phosphorylation : The process of ATP synthesis during oxidation of reduced coenzymes in ETC is called oxidative phosphorylation. Peter Mitchell (1961) proposed the chemiosmotic mechanism of ATP synthesis (Noble prize in 1978) which states that ATP synthesis occurs due to H+- flow through a membrane. It involves two steps :
(a) Development of proton gradient. At each step of ETC, the electron- acceptor has a higher electron -affinity than the electron-donor. The energy from electron-transport is used to move the proton (H+) from the mitochondrial matrix to inter-membranous or outer chamber. Three pairs of protons are pushed to outer chamber during the movement of electrons along route I while two pairs of protons are moved to outer chamber during the movement of electrons along route-II. This generates a pH-gradient across the inner mitochondrial membrane with protons (H+) concentration higher in the outer chamber than in the mitochondrial matrix. This difference in H+ concentration across the inner mitochondrial membrane is called proton-gradient(\[\Delta \]pH). Due to proton gradient, an electrical potential (Dy) is developed across the inner mitochondrial membrane as the matrix is now electronegative with respect to the intermembranous (outer) chamber. The proton gradient and membrane electric potential collectively called proton motive force.
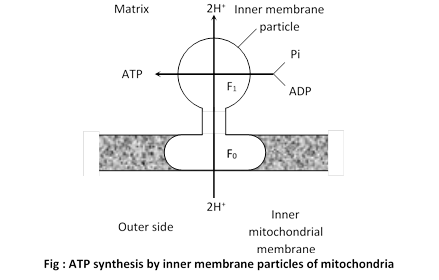
(b) Proton flow : Due to proton-gradient, the protons returns to the matrix while passing through proton channel of F0-F1 ATPase. This proton gradient activates the enzyme ATP synthetase or F0 - F1 ATPase
ATP synthetase controls the formation of ATP from ADP and inorganic phosphate in the presence of energy.
Important Tips
(i) Dinitrophenol (2,4-DNP) : It prevents synthesis of ATP from ADP because it directs electrons from CoQ to Q2.
(ii) Cyanide : It prevents flow of electrons from Cyt a3 to oxygen.
(iii) Carbon monoxide : It functions like cyanide.
(iv) Antimycin A: Transfer of electron from Cyt b to Cyt c1 is prevented.
(v) Rotenone : It checks flow of electrons from NADH /FADH2 to CoQ.
(i) 12.2 Kcal during transfer of electrons from NAD to FMN.
(ii) 15.2 Kcal during transfer of electrons from Cyt b to Cyt c.
(iii) 24.5 Kcal during transfer of electrons from Cyt a to Cyt a3.
(iv) Role of shuttle system in energy production : Glycolysis occurs in the cytoplasm outside the mitochondrion in which 2\[NAD{{H}_{2}}\] molecules are produced but ETC is located along inner mitochondrial membrane, so \[NAD{{H}_{2}}\]of glycolysis must enter inside the mitochondrion to release energy. But the inner mitochondrial membrane is impermeable to NADH2. In mitochondrial membrane, there are 2 shuttle-system, each formed of carrier-molecule.
These shuttle systems are :
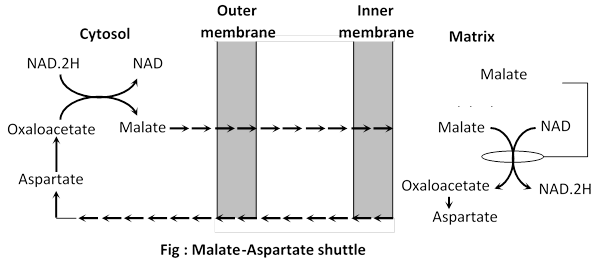
(a) Malate-Aspartate shuttle : It is more efficient and results in the transfer of electron from NAD. 2H in cytosol to NAD inside the mitochondrion, via NAD. 2H dehydrogenase as follows :
Electrons are transferred from NAD. 2H in cytosol to malate which traverses the inner mitochondrial membrane and reoxidised to form NAD. 2H thus resulting in the formation of oxaloacetate . Oxaloacetate does not readily cross the inner mitochondrial membrane and so a transamination reaction is needed to form aspartate which does traverse this barrier. As a result 3 ATP molecules are generated for each pair of electrons. Thus if this shuttle is predominant there is a gain of 38 ATP molecules by complete oxidation of one molecule of glucose.
(b) Glycerol-Phosphate shuttle : It is less efficient and results in the reduction of FAD inside the mitochondrion.
If this shuttle predominates the electrons from NAD. 2H are transferred to FAD inside the mitochondrion as follows. NAD. 2H reacts with dihydroxyacetone phosphate (DHAP) in cytosol to form glycerol phosphate which diffuses through outer mitochondrial membrane to the outer surface of inner membrane. There glycerol phosphate reacts with membrane dehydrogenase to form dihydroxyacetone phosphate (DHAP) which returns to cytosol. In this process FAD is reduced to FADH2. Electrons from FADH2 directly pass to Q and other components of ETC and results in the synthesis of 2 ATP for each molecule of FADH2. In this case complete oxidation of glucose will result in a gain of 36 ATP molecule.
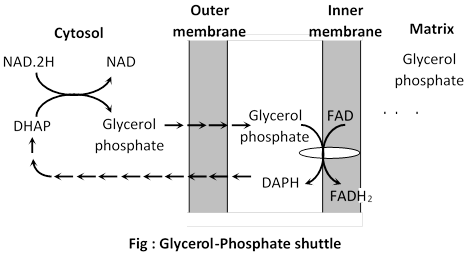
Which shuttle predominates depends on the particular species and tissues envolved, for example: 38 ATP are formed in kidney, heart and liver cell while 36 ATP molecules are formed in muscle cells and nerve cells. In these cells glycerol-phosphate shuttle is predominant and 2 ATP formed from NADH2.
Other pathways of glucose oxidation
(1) Entner-Doudoroff pathway
(i) Discovery: Entner-Doudoroff path discovered by Entner & Doudoroff. This pathway is also called glycolysis of bacteria.
Certain bacteria such as Pseudomonas sacchorophila, P. fluorescens, P. lindeneri and P. averoginosa lack phosphofructokinase enzyme. They cannot degrade glucose by glycolytic process.
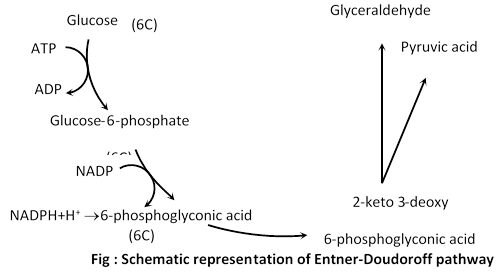
(ii) Description: In this pathway the glucose molecule first phosphorylated to Glucose-6-phosphate by ATP. Then it oxidised to 6-phosphogluconic acid by NADP which itself reduced to NADPH2 by the electrons released. The NADPH2 is channeled through ETS system to produce 3-molecules of ATP per NADPH2 molecule through ETS system to produce 3 molecules of ATP per NADH2 molecule and 1, 6-phosphogluconic acid is channeled to pyruvic acid. The main reaction are:
(a) \[\text{Glucose-6-phosphate}+\text{NADP }\to \text{6-Phosphogluconolactone }+\text{NADP}{{\text{H}}_{\text{2}}}\]
(b) 6-phosphoglyconolactone ® 2-Keto-3-deoxyphospho-6-gluconic acid
(c) 2-Keto-3-deoxyphosphogluconic acid ® Pyruvic acid+Glyceraldehyde-3-phosphate
(d) Glyceraldehyde-3-phosphate ® Pyruvic acid
The glyceraldehyde 3-phosphate by EMP pathway gets converted into pyruvic acid which can be further used up in the process.
(2) Pentose phosphate pathway
(i) Discovery : It is also called as Hexose monophosphate (HMP) shunt or Warburg Dickens pathway or direct oxidation pathway. It provides as alternative pathway for breakdown of glucose which is independent of EMP pathway (glycolysis) and Krebs cycle. Its existence was suggested for the first time by Warburg et al. (1935) and Dickens (1938). Most of the reaction of this cycle were described by Horecker et al.(1951) and Racker (1954).
(ii) Occurrence : Pentose phosphate pathway that exists in many organisms. This pathway takes place in the cytoplasm and requires oxygen for its entire operation.
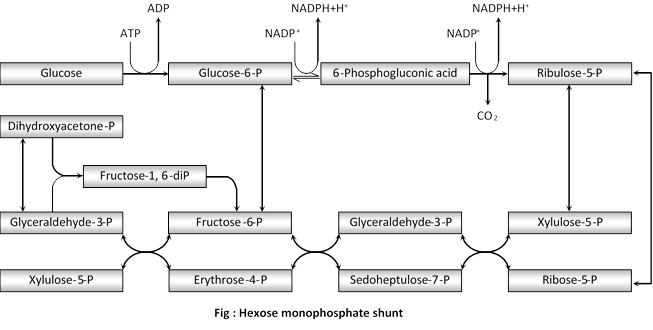
(iii) Description : There are two types of evidences is support of the existence of such an alternative pathway-works on the inhibiting action of malonic acid on the Krebs cycle and studies with the radioactive (C14).
Twelve molecules of NADH2 formed in the reaction can be oxidised back to 12 NADP with the help of the cytochrome system and oxygen of the air.
\[12\text{ NADP}{{\text{H}}_{\text{2}}}+6{{O}_{2}}\underset{\text{System}}{\mathop{\xrightarrow{\text{Cytochrome}}}}\,\text{12}{{\text{H}}_{\text{2}}}\text{O}+\text{12NADP}\]
In this electron transfer process, 36 molecules of ATP are synthesized.
The reaction can be summarised as follows :
\[\xrightarrow{\text{transketolase}}2\,\text{sedoheptulose}-\text{7phosphate}+\text{2 glyceraldehyde-3-phosphate }\]
\[\xrightarrow{\text{transketolase}}\text{2 fructose-6-phosphate}+\text{2 erythrose-4-phosphate}\]
\[\xrightarrow{\text{transketolase}}2\text{ fructose-6-phosphate}+\text{2 glyceraldehyde}\,\,\text{3-phosphate}\]
Sum total of the reaction :
\[6\text{ }Glucose-phosphate\text{ }+\text{ }12NADP+7{{H}_{2}}O5\text{ }Fructose-6-phosphate+6C{{O}_{2}}+\text{ }12NADP{{H}_{2}}+{{H}_{2}}P{{O}_{4}}\]
(iv) Significance of PPP
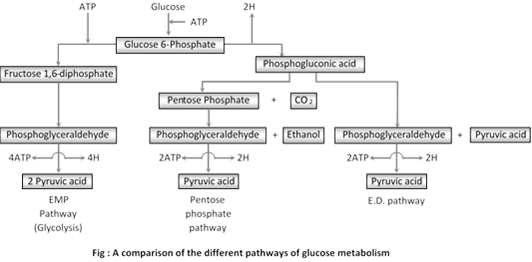
(3) Comparison of the different pathway of glucose metabolism
(4) Cyanide resistant pathway : Cyanide-resistant respiration seems to be widespread in higher plant tissues. Cyanide prevents flow of electron from Cyt a3 to oxygen, so called ETC inhibitor. In these plant tissues resistance is due to, a branch point in the ETS preceeding the highly cyanide-sensitive cytochromes. The tissues lacking this branch point, or alternate pathway and blockage of cytochromes by cyanide, inhibits the electron flow.
Significance
(a) The role of alternative pathway is that it may provide a means for the continued oxidation of NADH and operation of the tricarboxylic acid cycle, even through ATP may not be sufficiently drained off.
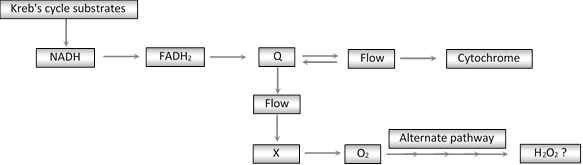
(b) It is significant in respiratory climateric of ripening fruits and leads to the production of hydrogen peroxide and super oxide, which in turn enhances the oxidation and breakdown of membranes.
(c) Necessary activities in the ripening process because peroxides are necessary for ethylene biosynthesis.




You need to login to perform this action.
You will be redirected in
3 sec

