Modern Concept Of Photosynthesis
Category : 11th Class
Photosynthesis is an oxidation reduction process in which water is oxidised to release O2 and CO2 is reduced to form starch and sugars.
Scientists have shown that photosynthesis is completed in two phases.
(1) Light phase or Photochemical reactions or Light dependent reactions or Hill's reactions : During this stage energy from sunlight is absorbed and converted to chemical energy which is stored in ATP and \[NADPH+{{H}^{+}}.\]
(2) Dark phase or Chemical dark reactions or Light independent reactions or Blackman reaction or Biosynthetic phase : During this stage carbohydrates are synthesized from carbon dioxide using the energy stored in the ATP and NADPH formed in the light dependent reactions.
Evidence for light and dark reactions in photosynthesis :
(1) Physical separation of chloroplast into grana and stroma fractions : It is now possible to separate grana and stroma fractions of chloroplast. If light is given to grana fraction in presence of suitable H-acceptor and in complete absence of \[C{{O}_{2}},\]then ATP and \[NADP{{H}_{2}}\]are produced (i.e., assimilatory powers). If these assimilatory powers (ATP and\[NADP{{H}_{2}}\]) are given to stroma fraction in presence of \[C{{O}_{2}}\]and absence of light, then carbohydrates are formed.
(2) Experiments with intermittent light or Discontinuous light : Rate of photosynthesis is faster in intermittent light (Alternate light and dark periods) than in continuous light. It is because light reaction is much faster than dark reaction, so in continuous light, there is accumulation of ATP and \[NADP{{H}_{2}}\]and hence reduction in rate of photosynthesis but in discontinuous light, ATP and \[NADP{{H}_{2}}\]formed in light are fully consumed during dark in reduction of \[C{{O}_{2}}\] to carbohydrates. Accumulation of \[NADP{{H}_{2}}\] and ATP is prevented because they are not produced during dark periods.
(3) Temperature coefficient studies : Blackman found that \[{{Q}_{10}}\] was greater than 2 in experiment when photosynthesis was rapid and that \[{{Q}_{10}}\] dropped from 2 often reaching unity, i.e., 1 when the rate of photosynthesis was low. These results show that in photosynthesis there is a dark reaction (\[{{Q}_{10}}\] more than 2) and a photochemical or light reaction (with \[{{Q}_{10}}\] being unity).
\[{{Q}_{10}}=\frac{\text{Reaction}\,\text{rate}\,\text{of}\,(t+10){}^\circ C}{\text{Reaction}\,\text{at}\,t{}^\circ C}\]
Light reaction (Photochemical reactions) : Light reaction occurs in grana fraction of chloroplast and in this reaction are included those activities, which are dependent on light. Assimilatory powers (ATP and\[NADP{{H}_{2}}\]) are mainly produced in this light reaction.
Robin Hill (1939) first of all showed that if chloroplasts extracted from leaves of Stellaria media and Lamium album are suspended in a test tube containing suitable electron acceptors, e.g., Potassium ferroxalate (Some plants require only this chemical) and potassium ferricyanide, oxygen is released due to photochemical splitting of water. Under these conditions, no \[C{{O}_{2}}\]was consumed and no carbohydrate was produced, but light-driven reduction of the electron acceptors was accompained, by \[{{O}_{2}}\] evolution.
\[\underset{\begin{smallmatrix} \text{Electron} \\\text{acceptor} \end{smallmatrix}}{\mathop{4F{{e}^{3+}}}}\,+\underset{\begin{smallmatrix} \text{Electron} \\\,\,\text{donor} \end{smallmatrix}}{\mathop{2{{H}_{2}}O}}\,\overset{\,\,\,\,\,\,\,}{\longleftrightarrow}\underset{\begin{smallmatrix} \text{Reduced} \\\,\text{Product}\end{smallmatrix}}{\mathop{4F{{e}^{2+}}}}\,+4{{H}^{+}}+{{O}_{2}}\uparrow \]
The splitting of water during photosynthesis is called photolysis. This reaction on the name of its discoverer is known as Hill reaction.
Hill reaction proves that
(1) In photosynthesis oxygen is released from water.
(2) Electrons for the reduction of \[C{{O}_{2}}\] are obtained from water [i.e., a reduced substance (hydrogen donor) is produced which later reduces \[C{{O}_{2}}].\]
Dichlorophenol indophenol is the dye used by Hill for his famous Hill reaction.
According to Arnon (1961), in this process light energy is converted to chemical energy. This energy is stored in ATP (this process of ATP formation in chloroplasts is known as photophosphorylation) and from electron acceptor NADP+, a substance found in all living beings NADP*H is formed as hydrogen donor. Formation of hydrogen donor NADPH from electron acceptor NADP+ is known as photoreduction or production of reducing power NADPH.
Light phase can be explained under the following headings :
(1) Transfer of energy : When photon of light energy falls on chlorophyll molecule, one of the electrons pair from ground or singlet state passes into higher energy level called excited singlet state. It comes back to hole of chlorophyll molecule within \[{{10}^{9}}\]seconds.
This light energy absorbed by chlorophyll molecule before coming back to ground state appears as radiation energy, while that coming back from excited singlet state is called fluorescence and is temperature independent. Sometimes the electron at excited singlet state gets its spin reversed because two electrons at the same energy level cannot stay; for some time it fails to return to its partner electron. As a result it gets trapped at a high energy level. Due to little loss of energy, it stays at comparatively lower energy level (Triplet state) from excited singlet state. Now at this moment, it can change its spin and from this triplet state, it comes back to ground state again losing excess of energy in the form of radiation. This type of loss of energy is called as phosphorescence.
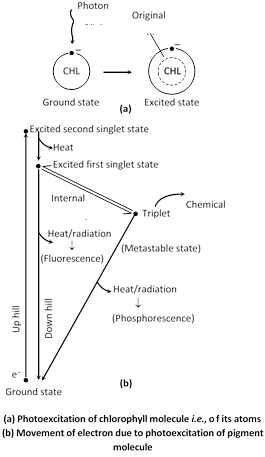
When electron is raised to higher energy level, it is called at second singlet state. It can lose its energy in the form of heat also. Migration of electron from excited singlet state to ground state along with the release of excess energy into radiation energy is of no importance to this process. Somehow when this excess energy is converted to chemical energy, it plays a definite constructive role in the process.
(2) Quantum yield
(i) Rate or yield of photosynthesis is measured in terms of quantum yield or \[{{O}_{2}}\] evolution, which may be defined as, "Number of \[{{O}_{2}}\]molecules evolved per quantum of light absorbed in photosynthesis."
(ii) Quantum requirement in photosynthesis = 8, i.e., 8 quanta of light are required to evolve one mol. of \[{{O}_{2}}.\]
(iii) Hence quantum yield \[=1/8=0.125\](i.e., a fraction of 1) as 12%.
(3) Emerson effect and Red drop : R. Emerson and C.M. Lewis (1943) observed that the quantum yield of photosynthesis decreases towards the far red end of the spectrum (680nm or longer). Quantum yield is the number of oxygen molecules evolved per light quantum absorbed. Since this decrease in quantum yield is observed at the far region or beyond red region of spectrum is called red drop.
Emerson et al. (1957) further observed that photosynthetic efficiency of light of 680nm or longer is increased if light of shorter wavelengths (Less than 680nm) is supplied simultaneously. When both short and long wavelengths were given together the quantum-yield of photosynthesis was greater than the total effect when both the wavelengths were given separately. This increase in photosynthetic efficiency (or quantum yield) is known as Emerson effect or Emerson enhancement effect.
\[E=\frac{\text{Quantum yield in combined beam}-\text{Quantum yield in red beam}}{\text{Quantum yield in far red beam}}\]
(4) Two pigment systems : The discovery of Emerson effect has clearly shown the existence of two distinct photochemical processes, which are believed to be associated with two different specific group of pigments.
(i) Pigment system I or Photosystem I : The important pigments of this system are chlorophyll a 670, chlorophyll a 683, chlorophyll a 695, \[{{P}_{700}}.\]Some physiologists also include carotenes and chlorophyll b in pigment system I. \[{{P}_{700}}\] acts as the reaction centre. Thus, this system absorbs both wavelengths shorter and longer than 680nm.
(ii) Pigment system II or photosystem II : The main pigments of this system are chlorophyll a 673, \[{{P}_{680}},\] chlorophyll b and phycobilins. This pigment system absorbs wavelengths shorter than 680nm only. \[{{P}_{680}}\] acts as the reaction centre.
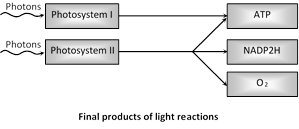
Pigment systems I and II are involved in non-cyclic electron transport, while pigment system I is involved only in cyclic electron transport. Photosystem I generates strong reductant NADPH. Photosystem II produces a strong oxidant that forms oxygen from water.
Comparison of photosystem I and photosystem II
|
S. No. |
Photosystem I |
Photosystem II |
|
(1) |
PS I lies on the outer surface of the thylakoids |
PS II lies on the inner surface of the thylakoid. |
|
(2) |
In this system molecular oxygen is not evolved. |
As the result of photolysis of water molecular oxygen is evolved. |
|
(3) |
Its reaction center is P700. |
Its reaction center is P680. |
|
(4) |
NADPH is formed in this reaction. |
NADPH is not formed in this reaction. |
|
(5) |
It participates both in cyclic and noncyclic photophosphorylation. |
It participates only in noncyclic photophosphorylation. |
|
(6) |
It receives electrons from photosystem II. |
It receives electrons from photolytic dissociation of water. |
|
(7) |
It is not related with photolysis of water. |
It is related with photolysis of water. |
(5) Photophosphorylation : Light phase includes the interaction of two pigment systems. PS I and PS II constitute various type of pigments. Arnon showed that during light reaction not only reduced NADP is formed and oxygen is evolved but ATP is also formed. This formation of high energy phosphates (ATP) is dependent on light hence called photophosphorylation.
Photophosphorylation is of two types :
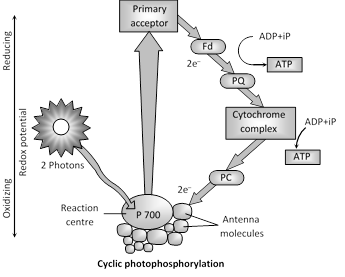
(i) Cyclic photophosphorylation : The system is found dominantly in bacteria. It involves only PS I. Flow of electron is cyclic. If NADP is not available then this process will occur. When the photons activate PS I, a pair of electrons are raised to a higher energy level. They are captured by primary acceptor which passes them on to ferredoxin, plastoquinone, cytochrome complex, plastocyanin and finally back to reaction centre of PS I i.e., P700. At each step of electron transfer, the electrons lose potential energy. Their trip down hill is caused by the transport chain to pump \[{{H}^{+}}\]across the thylakoid membrane. The proton gradient, thus established is responsible for forming (2 molecules) ATP. No reduction of NADP to \[NADPH+\text{ }{{H}^{+}}.\] ATP is synthesized at two steps.
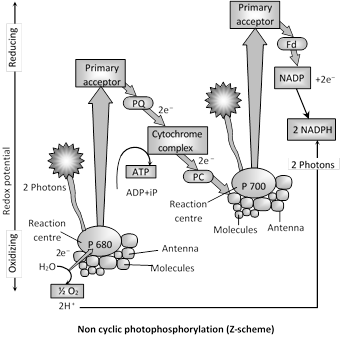
(ii) Non cyclic photophosphorylation : The system is dominant in green plants. It involves both PS-I and PS-II. Flow of electrons is unidirectional. Here electrons are not cycled back and are used in the reduction of NADP to \[NADP{{H}_{2}}.\]Here \[{{H}_{2}}O\] is utilized and \[{{O}_{2}}\] evolution occurs. In this chain high energy electrons released from \['P-680'\] do not return to \['P-680'\] but pass through pheophytin, plastoquinone, cytochrome \[{{b}_{6}}-f\]complex, plastocyanin and then enter \[P-700.\] In this transfer of electrons from plastoquinone (PQ) to cytochrome \[{{b}_{6}}-f\]complex, ATP is synthesized. Because in this process high energy electrons released from \['P-680'\] do not return to \['P-680'\] and ATP (1 molecules) is formed, this is called Noncyclic photophosphorylation. ATP is synthesized at only one step.
This non-cyclic photophosphorylation is also known as Z-scheme (because of shape of path of electron-flow) and this was given by Hill and Bendall (1960). Non-cyclic photophosphorylation or Z-scheme is inhibited by CMU and DCMU.
(DCMU is a herbicide which kills the weed by inhibiting \[C{{O}_{2}}\]fixation as it is a strong inhibitor of PS-II).
Pseudocyclic photophosphorylation : Arnon and his coworker (1954) demonstrated yet another kind of photophosphorylation. They observed that even in absence of \[C{{O}_{2}}\]and NADP, if chlorophyll molecules are illuminated, it can produce ATP from ADP and iP (Inorganic phosphate) in presence of FMN or vit. K and oxygen. The process is thus very simple and requires no net chemical change for the formation of ATP and water. Arnon called this oxygen dependent FMN catalysed photophosphorylation or pseudocyclic photophosphorylation which involves the reduction of FMN with the production of oxygen. FMN is an auto-oxidisable hydrogen acceptor with the effect that the reduced FMN is reoxidised by oxygen. Thus the process can continue repeatedly to produce ATP.
Since this process can be continuously self repeated, it appears that a single molecule of water should be sufficient to operate pseudocyclic photophosphorylation to meet the requirement of ATP.

Dark phase : The pathway by which all photosynthetic eukaryotic organisms ultimately incorporate \[C{{O}_{2}}\] into carbohydrate is known as carbon fixation or photosynthetic carbon reduction (PCR) cycle or dark reactions.
The dark reactions are sensitive to temperature changes, but are independent of light hence it is called dark reaction, however it depends upon the products of light reaction of photosynthesis, i.e., NADP .2H and ATP.
The carbon dioxide fixation takes place in the stroma of chloroplasts because it has enzymes essential for fixation of \[C{{O}_{2}}\]and synthesis of sugar.
The techniques used for studying different steps were Radioactive tracer technique using \[^{14}C\] (Half life – 5720 years), Chromatography and Autoradiography and the material used was Chlorella (Cloacal alga) and Scenedesmus (these are microscopic, unicellular algae and can be easily maintained in laboratory).
The assimilation and reduction of \[C{{O}_{2}}\] takes place in this reaction by which carbohydrate is synthesized through following three pathways :
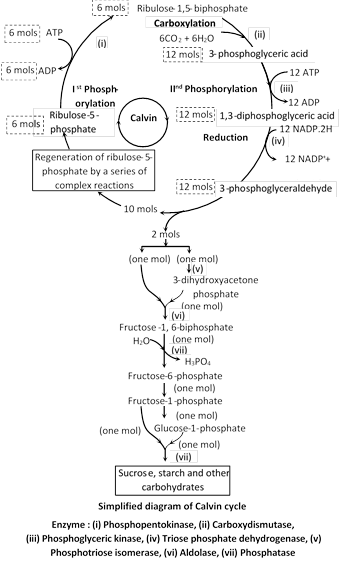
(1) Calvin cycle : Calvin and Benson discovered the path of carbon in this process. This is known as \[{{C}_{3}}\] cycle because \[C{{O}_{2}}\] reduction is cyclic process and first stable product in this cycle is a 3-C compound (i.e., 3-Phosphoglyceric acid or 3-PGA).
Calvin cycle is divided into three distinct phases : Carboxylation, Glycolytic reversal, regeneration of RuBP.
In this cycle, \[C{{O}_{2}}\] acceptor molecule is RuBP or RuDP (i.e., Ribulose 1, 5-biphosphate or Ribulose 1, 5-diphosphate). There occurs covalent bonding of \[C{{O}_{2}}\] to RuBP and the enzyme catalyzing this reaction is RuBP-carboxylase/oxygenase (Rubisco).
As calvin cycle takes in only one carbon (as\[C{{O}_{2}}\]) at a time, so it takes six turns of the cycle to produce a net gain of six carbons (i.e., hexose or glucose).
In this cycle, for formation of one mole of hexose sugar (Glucose), 18 ATP and \[12\text{ }NADP{{H}_{2}}\]are used.
The plants in which this pathway of \[C{{O}_{2}}\] reduction occurs, are called \[C-3\] plants.
About 85% of plant species are \[C-3\]plants, including cereals (e.g., barley, rice, oat, wheat), groundnut, sugarbeet, cotton, tobacco, spinach, soybean most trees and lawn grasses etc.
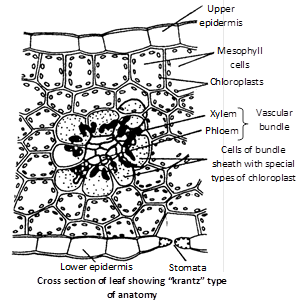
(2) Hatch and Slack cycle (C4 cycle) : Kortschak and Hart supplied \[C{{O}_{2}}\] to the leaves of sugarcane, they found that the first stable product is a four carbon \[({{C}_{4}})\] compound oxalo acetic acid instead of 3-carbon atom compound. The detailed study of this cycle has introduced by M.D. Hatch and C.R. Slack (1966). So it is called as "Hatch and Slack cycle". The stable product in \[{{C}_{4}}\] plant is a dicarboxylic substance. Hence it is called dicarboxylic acid cycle or DCA-cycle. \[{{C}_{4}}\] plants are true xerophytic plants.
The important \[{{C}_{4}}\] plants are sugarcane, maize, Sorghum, Cyperus rotundus, Digitaria brownii, Amaranthus, etc. These plants have "Kranz" (German term meaning halo or wreath) type of leaf anatomy. The vascular bundles, in \[{{C}_{4}}\] leaves are surrounded by a layer of bundle sheath cells that contain large number of chloroplasts. The chloroplasts in \[{{C}_{4}}\] leaves are dimorphic (Two morphologically distinct types). The chloroplasts of bundle sheath cells are larger in size and arranged centripetally. They contain starch grains but lack grana. The mesophyll cells, on the other hand, contain normal types of chloroplasts. Mesophyll and bundle sheath cells are connected by plasmodesmata. The mesophyll cells perform \[{{C}_{4}}\] cycle and the cells of bundle sheath perform \[{{C}_{3}}\] cycle.
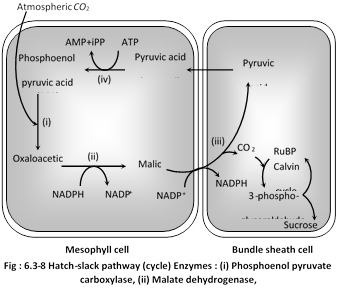
\[C{{O}_{2}}\]taken from the atmosphere is accepted by phosphoenolpyruvic acid (PEP) present in the chloroplasts of mesophyll cells of these leaves, leading to the formation of a \[4-C\]compound, oxaloacetic acid (OAA). This acid is converted to another 4-C acid, the malic acid which enters into the chloroplasts of bundle sheath cells and there undergoes oxidative decarboxylation yielding pyruvic acid (a \[3-C\]compound) and \[C{{O}_{2}}.C{{O}_{2}}\] released in bundle sheath cells reacts with Ribulose-1, 5-biphosphate (RuBP) already present in the chloroplasts of bundle sheath cells and thus Calvin cycle starts from here. Pyruvic acid re-enters mesophyll cells and regenerates phosphoenol pyruvic acid. \[C{{O}_{2}}\] after reacting with RuBP gives rise to sugars and other carbohydrates. In \[{{C}_{4}}\] plants, there are 2 carboxylation reactions, first in mesophyll chloroplast and second in bundle sheath chloroplast.
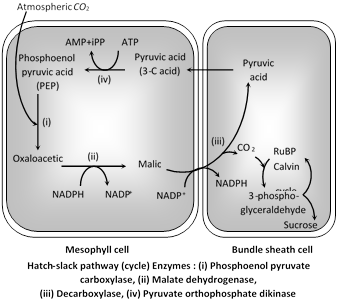
\[{{C}_{4}}\]plants are better photosynthesizers. There is no photorespiration in these plants. In \[{{C}_{4}}\] plants, for formation of one molecule of hexose (glucose) 30 ATP and \[12\text{ }NADP{{H}_{2}}\]are required.
Characteristics of \[{{\mathbf{C}}_{\mathbf{4}}}\]cycle
(1) \[{{C}_{\mathbf{4}}}\] species have greater rate of \[C{{O}_{2}}\]assimilation than \[{{C}_{3}}\] species. This is on account of the fact that
(i) PEP carboxylase has great affinity for \[C{{O}_{2}}.\]
(ii) \[{{C}_{4}}\] plants show little photorespiration as compared to \[{{C}_{3}}\]plants, resulting in higher production of dry matter.
(2) \[{{C}_{4}}\] plants are more adapted to environmental stresses than \[{{C}_{3}}\] plants.
(3) \[C{{O}_{2}}\]fixation by \[{{C}_{4}}\] plants requires more ATP than that by \[{{C}_{3}}\] plants. This additional ATP is needed for conversion of pyruvic acid to phosphoenol pyruvic acid and its transport.
(4) \[C{{O}_{2}}\]acceptor molecule in \[{{C}_{4}}\] plants is PEP. Further, PEP-carboxylase (PEPCO) is the key enzyme (RuBP-carboxylase enzyme is negligible or absent in mesophyll chloroplast, but is present in bundle sheath chloroplast).
(3) Crassulacean acid metabolism (CAM) : This dark \[C{{O}_{2}}\] fixation pathway proposed by Ting (1971). It operates in succulent or fleshy plants e.g., Cactus, Sedum, Kalanchoe, Opuntia, Agave, Orchid, Pineapple and Bryophyllum helping them to continue photosynthesis under extremely dry condition.
The stomata of succulent plants remain closed during day and open during night to avoid water loss (Scotactive stomata). They store \[C{{O}_{2}}\] during night in the form of malic acid in presence of enzyme PEP carboxylase. The \[C{{O}_{2}}\]stored during night is used in Calvin cycle during day time. Succulents refix \[C{{O}_{2}}\]released during respiration and use it during photosynthesis.
This diurnal change in acidity was first discovered in crassulacean plants e.g., Bryophyllum. So it is called as crassulacean acid metabolism.
Formation of malic acid during dark is called acidification or phase-I. Release of \[C{{O}_{2}}\]for actual photosynthesis during day is called deacidification or phase-II.

Characteristics of CAM pathway
(1) There is decrease in pH during the night and increase in pH during the day.
(2) CAM plants have enzymes of both \[{{C}_{3}}\] and \[{{C}_{4}}\] cycle in mesophyll cells. This metabolism enable CAM plants to survive under xeric habitats. These plants have also the capability of fixing the \[C{{O}_{2}}\]lost in respiration.
(3) Malic acid is stored in the vacuoles during the night which is decarboxylated to release \[C{{O}_{2}}\]during the day.



You need to login to perform this action.
You will be redirected in
3 sec

