Some Representative Pteridophytes
Category : 11th Class
![]()
Habitat
Selaginella is commonly called the little club moss or spike moss. Selaginella is mainly found in damp shaded places. A few species are xerophytic and can withstand the dry conditions for months together. In dry conditions, the plant rolls up into a compact ball and root system is disorganized. During the rainy conditions the ball on absorbing moisture, becomes green again. Such plants are called resurrection plants or bird's nest moss, e.g., S.lepidophylla and S. pilifera are xerophytes and sold in the market as novelties, S.bryopteris (Sanjeevani) and S.rupestris (ornamental).
The common epiphytic species are S. chrysocaulos, S. kraussiana, S. oregana, S. chrysorrhizos.
Structure
External structure : The plant body is sporophytic (2n), which is an evergreen and delicate herb having adventitious roots. The plants show great variation in their morphology. Some species are prostrate growing upon the surface (e.g., S.kraussiana), some are suberect (e.g., S.trachyphylla) and others are climbers (e.g., S.allegans). The stem is covered with four rows of small leaves, out of these two rows are of smaller leaves and two of large leaves species with dimorphic leaves such as S.kraussiana, S.helvetica, S.lepidophylla, S.chrysocaulos etc.
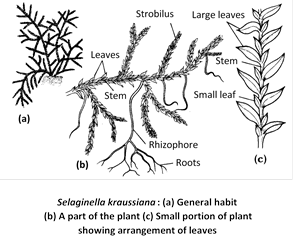
Leaves are sessile, ovate or lanceolate with acute apex. Unbranched midrib is present in the centre of each leaf. The leaves are ligulate, i.e., a flap-like outgrowth is present at the base on adaxial side called ligule. It may be fan-shaped or tongue-shaped or lobed or fringed. At the base of ligule, there is present a sheath of elongated cells called glossopodium (secretory).
The leaves possess a midrib but there is no venation. At the place of bifuraction of stem, a leafless, colourless, positively geotropic, elongated, cylindrical structure grows downwards. This is called the rhizophore and is quite different from the root in that it has no root cap.
Internal structure
Root : In root the stele is a protostele. It has a central core of xylem surrounded by phloem which is horse shoe shaped. It has a single protoxylem element (monarch). The xylem is exarch.
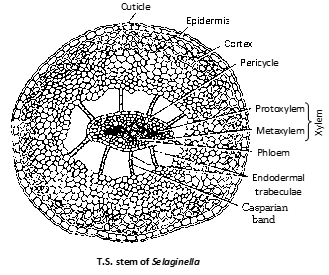
Stem : The stem is internally distinguishable into a single layered epidermis having no stomata. This is followed by cortex. The stele is suspended by unicelled (rarely multicelled) trabaculae (modified endodermal cell).
This layer, due to presence of casparian strips is regarded as endodermis. The stele is a protostele (haplostele) surrounded by a pericycle with a central core of xylem enclosed by phloem. Protostele is diarch and exarch. Stem of Selaginella kraussiana shows distelic condition.
Leaf : The leaf displays a simple structure. The mesophyll is uniform, being composed of elongated chlorenchymatous cells with large intercellular spaces. Each measophyll cells has one (S.martensii), two (S.kraussiana) or eight (S.willdenovii) chloroplasts. Each chloroplast has several pyrenoid-like bodies similar to Anthocerotales. The single midrib bundle is concentric, amphicribal (ectophloic) with annular or spiral tracheids surrounded by a few sieve elements.
Rhizophore : The anatomy of rhizophore is almost similar to root. The stele is a protostele. The xylem is exarch with several protoxylem groups. In S.kraussiana, centroxylic condition (having protoxylem in the centre surrounded by metaxylem elements) has been recorded.
Reproduction
Reproduction takes place by vegetative and sexual (by spores) method.
(1) Vegetative reproduction : It is of rare occurrence and may takes place by following methods :
Fragmentation : It occurs during very humid conditions. Some branches act as adventitious branches, which get separated from the plant and give rise to new Selaginella plants, e.g., in S.rupestris.
By resting buds : In some cases, terminal leaves get overlapped and become fleshy and form resting buds, which are means of vegetative reproduction, e.g., in S.chrysocaulos.
By tubers : In S.chrysorrhizos, some branches penetrate into substratum and at terminal ends swell to form tubers, which give rise to new plants.
By apogamy : In some cases, development of sporophyte occurs directly from gametophyte without intervention of sex organs, it is called apogamy and such plants are genetically haploid.
(2) Sexual reproduction : The reproductive structure in Selaginella is strobilus or spike. It is a sessile structure and develops at the terminal ends of the branches and its length varies from 1/4th of an inch to 2–3 inches in different species.
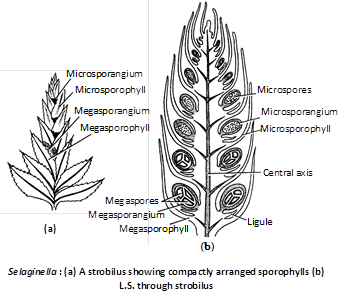
A strobilus is having many ligulate sporophylls arranged in cluster, each bearing a small, short, stalked sporangium on its upper surface. The sporangia are of two types :
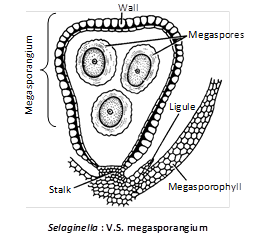
(i) Megasporangia : Borne on megasporophylls. Megasporangium is pale greenish and contains chalky white, yellow or orange megaspores.
The megasporangium is four-lobed structure with a 2-layered jacket, one layer of tapetum and a large number of microspore mother cell. However, only one megaspore mother cell is functional. After meiosis it produces 4 megaspores out of which 1–3 may degenerate. In S.rupestris, there is only a single megaspore.
(ii) Microsporangia : Borne on microsporophylls having a large number of small spores. Thus Selaginella is heterosporous. Microsporangium is pale yellow, oval or spherical body, with 2-layered jacket, one layered tapetum and a number of microspore mother cells which undergo meiosis and form haploid microspores. The main body consists of a wall having two layers, inside which are present numerous small microspores (400–2000). Development of sporangium is of eusporangiate type.
In most of the cases, the strobilus or spike bears two types of sporophylls; the lower are megasporophylls and the upper ones are microsporophylls. In S.kraussiana there is single megasporophyll at the base of spike and the rest of upper are microsporophylls.
In some cases strobilus contains either micro or megasporophyll, i.e., in S.gracilis and S.astrovirdis, while S. martensii and S.caulescens show intermixed micro and megasporophylls. Some sps. possess megasporangia on the ventral side and microsporangia on the dorsal side, e.g., S.oregana and S.inaequalifolia.

Mechanism of sporangial dehiscence : On maturation, the sporangium splits vertically from the upper end into two valves (vertical apical splitting). The lower cup-shaped portion shrinks and the spores come out through apical slits. This is brought about by cohesion owing to hygroscopic changes in the apical and lateral part of the sporangial wall. This liberation of spores takes place at intervals in small masses.
The spores starts germinating inside the sporangium before their release; this is known as precocious germination.
Germination of microspore : The microspore is a double layered structure and contains oil droplets. The outer wall exospore is much thicker (spiny) than inner endospore. It measures 15–50m in diameter. The microspore on germination forms the male gametophyte. The structure and development of male gametophyte was first described by Slagg (1932). The first division leads to formation of a small prothallial cell and a large antheridial cell.
The larger antheridial cell, by further divisions, gives rise to central group of four primary androgonial cells, surrounded by eight jacket cells. At this 13-celled stage (1 prothallial + 8 jacket cells and 4 primary androgonial cells), the microspore is shed from microsporangium. Each of the central groups of cells divides and redivides and finally forms about 256 spirally coiled antherozoids with two flagella (biflagellated); the jacket cells disintegrate. It takes about three weeks for germination of microspore and formation of antherozoids or sperms.
Germination of megaspore : The megaspore has three wall layers namely exospore, mesospore and endospore. It measures 1.5 – 5.0 mm in diameter. The megaspore on germination forms the female gametophyte. Generally the megaspore germinates inside the megasporangium (i.e., in situ). In some sps., megaspores are shed after the development of first archegonium, i.e., in S.kraussiana, while in S.apoda and S.rupestris, megaspores are not liberated till a well developed embryo is formed.
During the development of female gametophyte, the protoplasm after contraction forms a small sac-like structure. The outer wall bursts into two layers, the exospore and mesospore. At this stage, megaspore contains a haploid nucleus which by division produces many nuclei. Wall formation takes place in the upper beak-like portion and a small-celled cellular tissue is formed. This is one celled thick at the sides and three celled thick in the middle. This is female prothallus. Some superficial cells at apex enlarge and act as archegonial initials and form the archegonia. The megaspore bursts exposing the female prothallus. Vestigial rhizoids develop.
Archegonium are sessile and embedded type and consists of very short neck having a single neck canal cell and a venter, having a single ventral canal cell and an egg.
Fertilization
Usually the male gametophytes are shed from the microsporangium on the ground at 13-celled stage. Here they complete their development ultimately producing spermatozoids. These are liberated by the decay of the microspore wall. If the microspore falls near the mature female gametophyte, the sperms swim from the male gametophyte to reach archegonia and one sperm fuses with egg to form zygote. Water is necessary for fertilization and sperms are attracted due to malic acid.
Development of embryo or Sporophyte
The oosphere after fertilization gets surrounded by wall and become oospore. The oospore (zygote) divides transversely into two cells, the upper epibasal cell which forms suspensor cell and the hypobasal cell which develops into embryo.
The embryo differentiates into foot, root, primary stem with two rudimentary leaves and rhizophore. By growth of stem and the root, the young sporophyte becomes independent of the gametophyte tissue and falls on the ground where the primary rhizophore forms roots that grow into the soil and the plant starts independent life.
In some species of Selaginella, the archegonial initial develop apogamously into embryo. In S.intermedia, no microspores are formed. Here the embryo develops parthenogenetically from the egg. In S.helvetica, the archegonia fails to open and here also parthenogenetic development of embryo is seen.
There is distinct heteromorphic alternation of generations in Selaginella.
![]()
Habitat
Ferns live in moist, cool and shady places. They are perennial and evergreen. Fern Dryopteris filix-mas is commonly known as Beech fern or Male shield fern or Hay scented fern. There are about 150 sps. and 25 sps. have been reported in India. It is found in sub-tropical regions as well as warm temperate regions.
Structure
Fern plant is sporophytic (2n) with an underground rhizomatous stem, large aerial leaves or fronds and adventitious roots. Rhizome is sparingly branched in Dryopteris, moderately branched in Pteris and Adiantum (Adiantum is commonly called "Maiden hair fern" or walking fern because it propagates vegetatively by its leaf tips) and profusely branched in Pteridium. The younger leaf is called like a spring from apex downwards are called circinate venation.
Younger parts of leaves and rhizome are surrounded by brown hairy structures called scales or ramenta. Leaf bases are persistent.
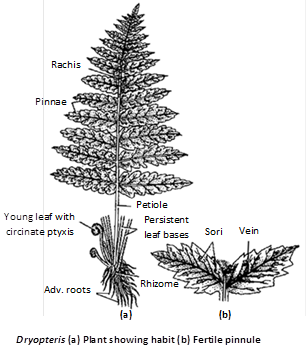
Root : In root the xylem is diarch and exarch with two phloem groups alternating the protoxylem.
Rhizome : Meristeles are found in fern rhizomes. Each meristele has its own endodermis followed by thin walled pericycle. It is amphicribal with mesarch xylem. The phloem lacks companion cells. In Dryopteris filix-mas the vasculature comprises a dictyostele consisting of a ring of meristele. In Pteris, however, the vasculature ranges from solenostele to polycyclic dictyostele.
Rachis : In the rachis of Dryopteris filix-mas there are 6–8 meristeles arragned in a horse-shoe like manner and single arched with hooked xylem in Pteris. The structure of the meristele is similar to that of rhizome.
Leaf lamina : Transverse section of lamina exhibits cuticularised upper and lower epidermis haivng chloroplast. The vascular strands lie embedded in mesophyll. Each strand is generally amphicribal but sometimes they are bicollateral also. The bundles in minor veins are collateral.
Reproduction
Vegetative reproduction : Vegetative reproduction can occur through fragmentation or rhizome and adventitious buds and these on separation gives rise to new fern plant.
Sexual reproduction : Sexual reproduction takes place through spores. Spores are born in sporangia. The spores are of one kind only (homosporous). When leaves are mature they bear groups of sori on the under surface of fertile pinnae. Such fronds are called sporophylls.
Sori are linear and submarginal in Pteris and Pteridium and median abaxial in Dryopteris. Each sorus is surrounded by a kidney-shaped covering called indusium. In Dryopteris, true indusium is present because this arises from placenta or placental tissue, from which sporangia arise. (In Pteris false indusium is there because it is formed by leaf margins).
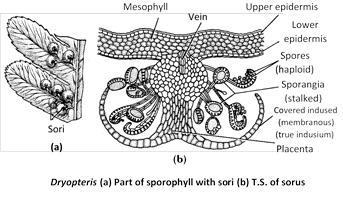
In sorus of Pteridium is covered by two flap like appendages that protect the sporangia. The upper flap is called false indusium and lower is called the true indusium. In the centre of sorus, the vein ends into placental tissue from where arise a number of sporangia. The sorus is mixed in Dryopteris (i.e., no definite arrangement of sporangia).
Sporangium : The sporangial development is leptosporangiate i.e., it develops from a single superficial initial. (In eusporangiate type the sporangium arises from a group of initials). A sporangium is distinguishable into a stalk and a capsule. The stalk is multicelled and biseriate.
The capsule is oval or elliptical and biconvex in shape. It consists of a single layered wall followed by double layered tapetum that encloses the archesporium. The archesporial cells divide and redivide to form a mass of sporogenous tissue. Most of the sporogenous cells behave as spore mother cells. They undergo meiosis to form tetrahedral tetrads of (haploid) spores. As a result 32–64 spores are formed in each capsule. The tapetal layer is nutritive. It degenerates at maturity of the sporangium.
A row of specially thickened cells, known as annulus partially surrounds the capsule. At one side of the annulus, there is a stomium composed of thin walled cells, through which capsule dehisces. The annulus and stomium help in the dehiscence of sporangium and ultimately in the release of spores.
Gametophytic generation
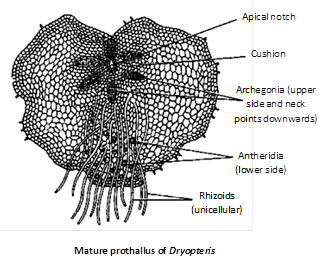
Spores : It is the first cell of gametophytic generation. Spores are double layered. The outer wall exospore is much thicker than inner endospore. On approach of favourable condition spore germinates to form a filamentous gametophyte which develops into green and heart shaped prothallus.
Prothallus : Fern prothallus is monoecious but protandrous (antheridia mature first). Antheridia are present in between the rhizoids while archegonia are present near the apical notch.
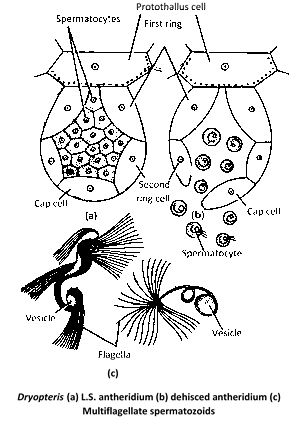
Antheridium : It consists of a three celled jacket enclosing a mass of androgonial cells. The two lower jacket cells are ring like (first and second ring cells), and the terminal cell is called as opercular cell or cover cell or cap cell. Sometimes, there may be two cap cells and in that case the jacket is four celled. The last generation of androgonial cells forms the androcytes. There may be 20–25 androcytes in an antheridium, each of which is metamorphose into a spirally coiled, multiflagellate antherozoid.
Archegonium : It is a flask-shaped structure having venter and neck. Neck is projected out of the prothallus and is curved posteriorly.
Venter is having basal large egg cell and upper small venter canal cell. The neck is having single neck canal cell but is binucleate. Venter is not having any covering or jacket but neck is surrounded by jacket of 4 vertical rows of cells.
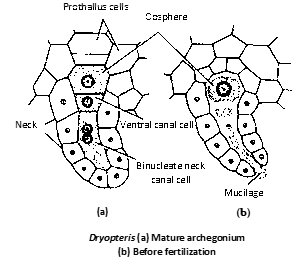
Fertilization and development of sporophyte : Fertilization takes place in the presence of water. The antherozoids are attracted towards archegonium due to the presence of malic acid in the mucilage oozing out from archegonia.
A single antherozoid is able to fuse with egg to form zygote (2n), which is beginning of sporophytic generation.
Zygote divides first by vertical division, followed by another vertical division and quadrat stage is formed. Then octant stage is formed by transverse division.
The foot and root develop from four hypobasal cells and cotyledons as well as stem develop from epibasal cells and thus sporophytic plant is formed. At maturity foot is hemispherical mass of gametophyte from which it absorbs food.
Generally a single sporophytic plant develops from single gametophyte or prothallus. The fern sporophyte is initially dependent upon gametophyte but later on becomes independent.
The life cycle is diplohaplontic with heteromorphic alternation of generation.



You need to login to perform this action.
You will be redirected in
3 sec

