Some Representative Rymnosperms
Category : 11th Class
![]()
Habitat
It is commonly known as pine with about 90 species among which six species are found in India. (N. East and N. West Himalayas) occurring in wild state. These are Pinus gerardiana (Chilgoza pine), P. Wallichiana (Blue pine or Kail), P.roxburghii (Chir pine), P.merkusii (Teenasserim pine), P.insularis (Khasi pine), and P.armandi (Armand's pine). In addition to these, 4 sps. of exotic pines, i.e., P.montana, P.laricia and P.sylvestris (Scotch pine) and P.strobus (white pine) have been introduced in India. P.excelsa are found at maximum height i.e., grow upto 3500 m above see level.
Structure
External structure : Pinus is an evergreen, perennial plant of xerophytic nature. Mostly the species are tall and straight. The whorled branching gives a typical conical or excurrent appearance to the plant (due to apical dominance). The plant body is sporophyte and the plants are monoecious. The plant body is differentiated into roots, stem and leaves.
Root : A prominent tap root is present which does not penetrate deep into the soil. Lateral roots which develops later, grow extensively and help in anchoring the plant in the soil. Root hairs are scanty. Ectotrophic mycorrhiza i.e., symbiotic association of some fungal hyphae with the ultimate branches of roots, is of common occurrence.
Stem : The stem is erect, thick, cylindrical and branched. The branching is monopodial type. The main stem is covered by scaly bark. Branches are developed from the buds present in the axil of scale leaves and appear to be in whorls. These branches develop every year and help in calculating the age of the plant.

Branches are of two types :
(1) Long shoots or Branches of unlimited growth : These have apical buds, grow indefinitely in whorls each year from the buds in the axil of scale leaves. These shoots spread out horizontally and bear scale leaves on them.
(2) Dwarf shoots or Branches of limited growth : These branches lack apical buds and grow for a definite or short period. They arise in the axil of scale leaves on long shoots.
Leaves : The leaves are of two types i.e., dimorphic – scale leaves and foliage leaves.
Scale leaves : The scale leaves are small membranous and brown. They are present on both types of branches (i.e., long and dwarf shoots). Scale leaves are non-photosynthetic. These protect the young buds.
Foliage leaves : The foliage leaves are green, needle like and are born at the tips of the dwarf shoots only. Their size and number is different in different species. The dwarf shoot with needles is called a spur. On the basis of number of needles, spur is of different types as :
Monofoliar (with one needle), e.g., P. monophylla.
Bifoliar (with two needles), e.g., P. merkusii and P. sylvestris.
Trifoliar (with three needles), e.g. P. gerardiana and P. roxburghii.
Pentafoliar (with five needles), e.g., P. wallichiana, P. excelsa.
Internal structure
(1) Root : The young root of Pinus is identical with the dicot root. A T.S. of root reveals the following structures.
Epiblema : It is the outermost layer of compactly arranged cells. It gives out many thin and unicellular root hair.
Cortex : It is composed of many layeres of thin walled parenchymatous cells.
Endoermis : A single layer of suberized cells.
Pericycle : Endodermis is followed by multilayered pericycle.
Vascular tissues : Radial vascular bundles are present.
Xylem : Exarch condition with bifurcated (Y-shaped) protoxylem. Resin canal is present between two arms. Xylem is devoid of vessels.
Phloem : Alternating with the xylem groups are present phloem patches. Companion cells are absent.
Pith : Pith is generally absent. If present, it is very small and made-up of parenchymatous cells.
Secondary growth : In young roots, cambium is absent but at maturity below the phloem patches, arches of cambium are formed. It cuts off secondary xylem on the inner side and secondary phloem on the outer side.
The cells of the outermost layer of pericycle form cork cambium (phellogen), which cuts off phellem (cork) on the outer side and phelloderm (secondary cortex) on the inner side. Finally epiblema ruptures and the cork layer is exposed.
(2) Stem : T.S. of a young shoot shows following tissue.
Epidermis : It is the outermost layer made up of small compactly arranged cells (heavily cuticularised).
Hypodermis : Below epidermis are 4–5 layers of sclerenchymatous cells constituting hypodermis.
Cortex : Inner to the hypodermis is a wide zone of cortex, some cells are filled with tannin.
Endodermis : It is the innermost layer of the cortex, made-up of single layer of cells.
Vascular cylinder (Stele) : It is of eustelic type having a ring of 5–8 closely arranged vascular bundles. Vascular bundles are conjoint, collateral and open.
Xylem : It is endarch, consists of only tracheidal cells, vessels are absent. Therefore wood is known as non-porous.
Protoxylem consists of annular and spiral tracheids. Metaxylem tracheids have uniseriate bordered pits on their radial walls. These are also having bars of sanio.
Phloem : It is situated on the outer side of vascular bundle and is made-up of phloem parenchyma and sieve cells. Companion cells are lacking.
Cambium : In between the xylem and phloem of each vascular bundle, there is a strip of intrafascicular cambium.
Medullary ray : In between the vascular bundles is a zone of parenchymatous cells connecting the pith and the cortex.
Pith : In the centre of the stem is a zone of thin-walled parenchymatous cells known as pith. Some of the pith cells are filled with resinous substances.
Secondary growth : Secondary growth is similar to that of dicot stem. Wood is pycnoxylic and monoxylic. Vascular rays are linear (uniseriate) but fusiform (multiseriate) area of passage of resin ducts. These cambium cells cut cells on the inner side forming secondary xylem and on outer side secondary phloem.
The ring of primary cambium remains active only for a year. The activity of the cambium stops in the winter season and again resumes in the following spring. The secondary xylem thus formed clearly shows a number of annual rings. Each annual ring consists of a zone of spring wood and autumn wood.
Autumn wood : It is formed during autumn season and the cells of this wood are smaller, squarish and thick.
Spring wood : It is formed during spring season. The cells of this wood are thinner, large and polygonal. The wood is termed as pycnoxylic (compact and hard).
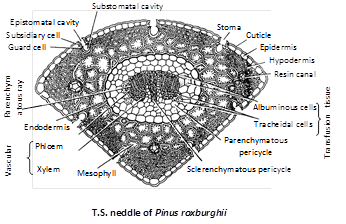
(3) Leaf (Needle) : The outline of foliage leaf varies according to the number of needles in the spur, i.e., in monofoliar spur of P. monophylla, it is circular, in bifoliar spur of P.sylvestris, it is semicircular and is triangular in trifolial spur of P. roxburghii. Internal structure of the needle is same in all species of Pinus. Needle shows xerophytic characters.
Epidermis : It has a single-layered, thick-walled epidermis, covered with thick cuticle and is interrupted by sunken stomata throughout the surface (amphistomatic).
Stomata : Each stoma has two guard cells and two subsidiary cells. It opens outside into a cavity called vestibule and inside into a substomatal cavity.
Hypodermis : Below the epidermis is present a few layered thick sclerenchymatous hypodermis. It helps in mechanical support.
Mesophyll : There is no differentiation into palisade and spongy parenchyma. The cells of this region are thin-walled, parenchymatous, polygonal, compactly arranged, having chloroplasts and starch grains. Peg-like infoldings arise from the inner surface.
Vascular cylinder : It is surrounded by single-layered endodermis having barrel-shaped cells with casparian strips.
Pericycle : Just below the endodermis is multilayered pericycle having a T-shaped mass of sclerenchymatous cells between two vascular bundles. Transfusion tissue occurs on the side. Each bundle is collateral, open and endarch.
The needle of P.monophylla has a single vascular bundle whereas in P.roxburghii, the number is two.
Reproduction
Pinus reproduces only by means of spores. Unlike Cycas, here the micro and megasporophylls form compact male and female cone or strobilus respectively.
Pinus plant is sporophyte (2x), heterosporous (producing two type of spores – microspore and megaspore), monoecious (male and female cones are borne on same plant) and autoicious (male and female cones are borne on different branches).
(1) Male cone or Staminate strobilus : The male cones are borne in a cluster on a branch of unlimited growth behind the apical bud, in the axil of a scale leaf. A male cone is, thus, equivalent to a dwarf shoot.
In a cluster, there may be 15–140 male cones. The male strobilus is an ovoid structure measuring 2 to 4 cm. in length and 0.5 to 0.7 cm. in diameter. A cone consists of a central axis bearing 60–135 microsporophylls in spiral manner. It is, therefore, comparable to male flower of angiosperms.
Microsporophyll : The microsporophylls or 'stamens' are spirally arranged in a compact manner on the cone axis. The microsporophyll is a brown coloured triangular structure consisting of a short stalk or 'filament' and a leaf like flattened structure called 'anther'.
Each sporophyll is provided with two microsporangia on its abaxial surface. The terminal sterile portion of the sporophyll is turned upward to protect the upper sporangia. It is called apophysis. Some of the lower microsporophylls are sterile having no sporangia associated with them.
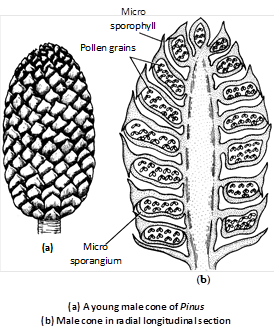
Microsporangium : The microsporangia are sessile elongated, cylindrical, structures. The sporangial development is of eusporangiate type. Each sporangium consists of a 2–3 layered wall. The inner most wall layer is called tapetum, which encloses a mass of sporogenous tissue. The sporogenous cells divide and re-divide and finally behave as microspore mother cells or pollen mother cells (PMC). The PMC undergo meiosis to form tetrahedral tetrads of microspores. The tapetum is a nutritive layer which degenerates at maturity of the anther.
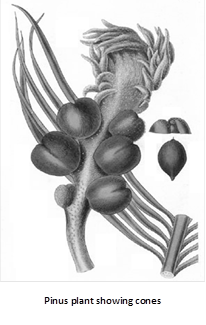
(2) Female cone (ovulate strobilus) : The female cone is an elongated, ovoid structure comprising a central cone axis on which the ovuliferous scales and bracts are spirally arranged in acropetal order. Usually the cone is 15 – 20 cm. long but in P. lambertiana they are 60 cm. long. The female cones take three years time to develop and mature.
The cones are produced in clusters of 1 to 4 from places where normally dwarf or spur shoots have developed. They arise in a group of 1 – 4 cones on a long shoot in the axil of a scale leaf in place of a dwarf shoot. In the first year, the female cone is reddish-green measuring about 1–2 cm in length having compactly arranged sporophylls. The second year cone is much larger, again with compact sporophylls. In the third year, the cone axis elongated and hence the sporophylls separate from each other.
Megasporophyll : Each megasporophyll is differentiated into two parts – Lower part is bract scale and upper part is ovuliferous scale.
Bract scales : These are small, dry, membranous structures attached with the cone axis directly. These are also known as carpellary or cover scales.
Ovuliferous scale : This is a woody, brownish structure borne on the dorsal side of the bract scale. Each ovuliferous scale is triangular with narrow basal part and upper broader part in the form of disc, known as apophysis. The apophysis appears to be rhomboidal and possesses a small point known as umbo.
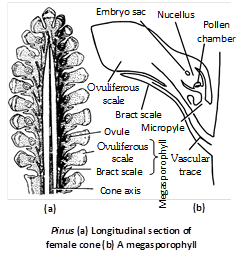
Megasporangium : Each ovule is an oval and anatropous structure consisting of a central mass of parenchymatous tissue, the nucellus, surrounded by a two lipped protective covering the integument which is united with nucellus except at the micropylar end where it prolongs to form a short tube beyond the nucellus. A small space is left in the upper region of nucellus below the integument, which is known as pollen chamber. Integument is differentiated into 3 layers although differentiation is not so distinct as in Cycas.
Outer fleshy layer : Made up of thin walled cells which disappears at maturity.
Middle stony layer : Very conspicious.
Inner fleshy layer : Inner fleshy layer is well developed.
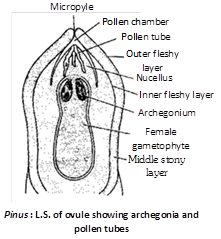
At the apex of the nucellus, a hypodermal cell gets enlarged and differentiated, it is called archesporial cell. The archesporial cell divides periclinally into an upper tapetal cell which forms tapetum, the nourishing layer, and the lower megaspore mother cell. This megaspore mother cell (sporogenous cell) divides reductionally to form a linear tetrad of haploid megaspores. Out of the four megaspores, three lying towards the micropyle degenerate. The chalazal one matures into a functional megaspore.
The gametophyte : The sporogenesis results in the formation of micro and megaspores representing the first gametophyte cells. They undergo gametogenesis so as to form the male and female gametophytes respectively.
Male gametophyte : The unicelled microspore undergoes three divisions of mircogametogenesis, so as to form a four-celled pollen grain or microgametophyte or male gametophyte.
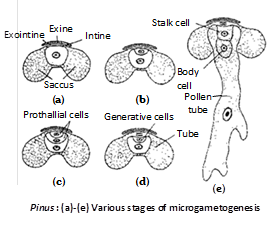
There are two prothallial cells, a generative cell and a tube cell. The pollen grains, at maturity are protected by three wall layers. The outermost wall layer, called exine or cappa is cutinised. the second wall layer is called exointine or capulla. It forms two balloon like outgrowths, on either side, called wings or saccus. The third wall layer is thin and called intine or tenuitas. A maturity the microsporangia dehisce by a longitudinal slit and the pollen grains are dispersed at 4 – celled stage. Since, a large number of grains are set free from a cluster of male cones in the form of pale-yellow cloud the phenomenon is often described as shower of sulphur or shower of golden dust.
Female gametophyte : The functional megaspore enlarges. A vacuole develops in the centre and then its nucleus divides freely to form about 2000 nuclei. Initially, multinucleate tube like cells are formed called alveoli. Later, wall formation starts from periphery and proceeds towards the centre. As a result, cellular female gametophyte or female prothallus or megagametophyte or endosperm is formed. The cells of the nucellus surrounding the female gametophyte now get modified and form a nutritive layer called endosperm jacket or spongy layer. The 'endosperm' of Pinus is a haploid gametophytic tissue formed before fertilization.
Archegonium : Near the micropylar end, one to five archegonia are differentiated in the prothallus. Each archegonium at maturity consists of eight neck cells arranged in two tiers of four cells each and a venter having a small ventral canal cell and a large egg. The ventral canal cell disorganizes before fertilization. Neck canal cells are absent.
Pollination : The pollination in Pinus is anemophilous. The wings of pollen grains are helpful in pollination. Just before pollination the female cone axis elongates separating megasporophyll from each other. This fascilitates pollen grains to reach ovules. There is a long interval of about a year between pollination and fertilization.
Post pollination changes in the male gametophyte : The exine ruptures and the intine protrudes out to from the pollen tube that grows through the nucellar tissue. Simultaneously, the generative cell divides to form a stalk cell and body cell. The body cell then divides to form two male gametes, which are non-flagellate.
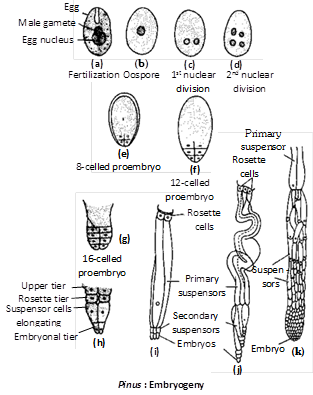
Fertilization : The mode of fertilization was discovered by Goroschankin (1883). After reaching the neck of the archegonium, the tip of the pollen tube ruptures releasing the two male gametes. The ventral canal cell degenerates and the neck cell split apart. Out of the two, one male gamete fuses with the egg to form the zygote. The second male gamete along with the stalk and body nuclei disintegrate.
Embryogeny : The proembryonal development in Pinus was studied by Buchholz (1918). The zygotic nucleus moves toward the base and then divides to form four nuclei. These nuclei organise into four quadrately arranged (diagonally opposite) cells with open upper end. The four cells divide simultaneously thrice to form four tiers of four cells each. These tiers are designated from top downwards as open tier, rosette tier, suspensor tier and apical tier. Since only a part of the oospore is involved in the formation of the embryo, the development is said to be meroblastic.
The cells of suspensor tier elongate pushing the embryonal cells into the 'endosperm'. The four suspensor cells due to considerable elongation may become coiled. These cells may divide transversely to form secondary suspensor or embryonal tubes.
By another transverse division, two whorls of embryonal tubes, designated as first and second series of embryonal tubes, are formed. All the four cells of embryonal tier separate from one another and develop into four independent embryos. The formation of more than one embryos from one oospore is called cleavage polyembryony.
Another type of polyembryony found in Pinus is simple polyembryony i.e., when more than one embroys are developed as a result of fertilization of different archegonia. Thus in Pinus although both types of polyembryony are found but at maturity seed contains only one embryo as food is not sufficient for survival of many embryos. The embryo soon gets differentiated into radicle, plumule, hypocotyl and cotyledons. The number of cotyledons is always more than two (Schizocotyly).
Seed formation : Seed of Pinus is winged. The wing develops from the upper surface of ovuliferous scale. Seed has thin with hard outer coat, which is peeled off, a stony coat, papery coat, cap like perisperm and food laden endosperm which encloses a central embryo. Embryo possesses 9–14 cotyledons (P.roxburghii). A seed represents three generations – parents sporophyte (testa, tegmen and perisperm, if present), new sporophyte (embryo) and female gametophyte or endosperm.
Seed germination : The seeds may remain dormant for several years. The germination of seed occurs when the environmental conditions are favourable. The radicle protrudes out through the micropyle and enters the soil forming the primary root. The plumule comes out and along with cotyledons it is pushed in air due to elongation of hypocotyl. The germination is, therefore, epigeal. The plumule forms a few juvenile leaves or prophylls. The juvenile leaves are spirally arranged on the branch of unlimited growth. Long shoots arise in their axis. Later on, they dry up as scales. The rate of growth of Pinus is quite slow.
Economic importance
(1) Seeds of some species are edible e.g., P. gerardiana (chilgoza), P. edulis.
(2) Fossilized resin (amber) is obtained from P. succinifera and is of great commercial value.
(3) Some species of Pinus are cheap source of cellulose.
(4) Some species are used for manufacture of paper.
![]()
Habitat
Cycas is an evergreen palm-like plant. It is the only genus of family Cycadaceae represented in India. Cycas has approximately 20 species found in Australia, New Zealand, Japan, China, India, Burma (Myanmar) and Pacific Islands.
In India, four Cycas species are common in Orissa, Bengal, Assam, Tamilnadu, Karnataka and Andaman.
(1) Cycas revoluta : It is a native of China and Japan and is locally called Tesso. In our country, it is called 'Sagopalm'. Due to its primitive characters, it is also called living fossil. It is upto 10 ft tall.
(2) Cycas circinalis : Plants are about 12 to 15 ft tall and distributed upto 3500 ft. In Hindi, it is called as Janglimadan mast-ka-phul.
(3) Cycas rumphii : Plants are about 12 ft tall. It is also cultivated in Indian gardens. In Tamil, it is called Kama, Paiyindu.
(4) Cycas beddomei : It is found in Trimula-hills of Andhra Pradesh.
Structure
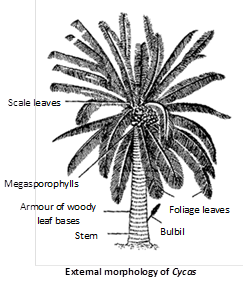
External structure : It looks very much like a palm tree or a tree fern, hence also called palm fern. Fully grown plants attain a height of 2– 5 m although C. media attains a height of 20m. The main plant body is differentiated into root, stems and leaves.
(1) Roots : Roots arise from lower part of stem and are of two types :
(i) Normal roots : These form a primary tap root system. These are positively geotropic and their main function is to absorb water and nutrients.
(ii) Coralloid roots : From the lateral branches of the normal roots are formed dichotomously branched, apogeotropic, bluish green coralloid roots. Anabaena cycadacearum, Nostoc and bacteria are found in their cortex. It is an example of symbiosis. It helps in fixation and absorption of nitrogen. As these roots possess lenticels, they also help in respiration.
(2) Stem : Stem is thick, cylindrical, columnar, small, aerial and unbranched. It is covered with persistent leaf bases and scale leaves, which are found in alternate whorls. There is a crown of foliage leaves at the apex of the plant.
(3) Leaves : Cycas has two types of leaves (dimorphism).
(i) Scale leaves : These are reduced form of foliage leaves without lamella and are arranged in a compact spiral and alternate manner around the apex and bears no reproductive structures. These are protective in nature. A single scale leaf is a brown, dry, woody, triangular structure, covered with brown hairs or ramenta.
(ii) Foliage leaves : These are green unpinnately compound present on the apex of the plant forming a crown. These leaves are upto 3 metres in C. circinalis. Leaves are leathery and thick, some leaflets at the base of the rachis are reduced to spines. These are mainly photosynthetic in nature. Leaves in Cycas show xerophytic characters.
Internal structure
(1) Root
Normal root : The structures of normal root resembles dicotyledonous root. T.S. of normal root reveals the following structures.
Epiblema : This is the outermost layer with unicelled root hairs.
Cortex : Just below the epiblema is multilayered parenchymatous cortex. Some tannin cells are present in the cortex.
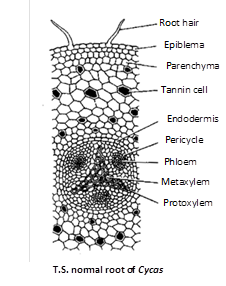
[Endodermis : Below the cortex is present endodermis which is made up of barrel-shaped cells and below it is a layer of pericycle.
Vascular tissues : It consists of xylem and phloem which are radially arranged, i.e., on different radii.
Pith : It is generally absent.
Secondary growth : It is like dicotyledonous plants.
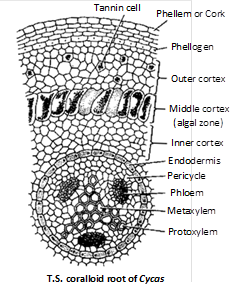
Coralloid root : Structure of stele is similar to normal roots but cortex is divided into three zones :
Outer cortex : Having several layers of parenchymatous cells.
Middle cortex (Algal zone) : Filled with blue green algae, Anabaena and Nostoc.
Inner cortex : Having several layers of parenchymatous cells. Roots are diarch, triarch and sometimes polyarch.
(2) Stem : It resembles a dicotyledonous stem having the following tissues :
Epidermis : It is the outermost incomplete layer ruptured due to persistent leaf bases. It is made up of compactly arranged thick-walled cells.
Cortex : Cortex is large, thin-walled, parenchymatous, having a number of mucilage canals. Starch grains are found in the cortex.
Endodermis and pericycle : These layers are not very clear.
Stele : Vascular cylinder is very small having numerous small closely arranged vascular bundles, which are conjoint, collateral and open. Xylem is endarch and consists of tracheids, which have spiral thickening in protoxylem and scalariform thickenings in metaxylem. Phloem is devoid of companion cells. Albuminous cells are found in phloem.
Leaf traces : There are several leaf traces present in the cortex. Four vascular bundles enter the base of leaf, two of these are direct and other two arise from the stele of opposite side and after making semicircle, they enter the leaf. These indirect leaf traces are known as girdling leaf traces or leaf girdles.
Pith : It is large, parenchymatous and is having a number of mucilage canals. Starch grains are also found in pith.
Secondary growth : The secondary growth in initiated by the formation of a cambium ring due to the development of interfascicular cambium strips and their subsequent joining with the intrafascicular cambium. This ring cuts secondary xylem on the inner side and secondary phloem on the outer side in addition to secondary medullary rays on both sides. This cambium ring now ceases to function another cambium now arises from pericycle or inner layers of cortex.
The new cambium functions in the normal way like the old one. Thus, concentric rings of secondary xylem and secondary phloem are formed. Such a wood is called as polyxylic i.e., comprising more than one xylem cylinders. Due to the presence of alternating rings of thin walled tissue (phloem) the wood of xylem remains loose and hence it is described as manoxylic. The growth in the extrastelar region takes place by the formation of a phellogen (cork cambium) which cuts off phellem (cork) on the outer side and phelloderm (secondary cortex) on the inner side. The three layers jointly constitute the periderm. The secondary growth pattern of Cycas resembles some dicots showing abnormal secondary growth. Secondary wood is devoid of vessels.
The secondary xylem is made up of tracheids showing multiseriate bordered pits. Bars of sanio have been observed by Sifton, 1915 in the tracheids of C. revoluta. The secondary phloem comprises sieve cells and fibers.
(3) Leaf
(i) Rachis : The cross section of the rachis is almost circular with two depressions on upper lateral sides where the leaflets are attached.
Epidermis : The outermost layer is epidermis with thick cuticle having stomata.
Hypodermis : Epidermis is followed by a well developed hypodermis, differentiated into outer chlorenchymatous and inner sclerenchymatous regions.
Ground tissue : Below the hypodermis is well developed parenchymatous ground tissue with mucilage canals. The vascular bundles are arranged forming an inverted omega (W). Each vascular bundle is surrounded by a sclerenchymatous sheath and is conjoint, collateral and open. In most parts of the rachis, xylem is mesarch, i.e., centripetal xylem towards periphery and two patches of centrifugal xylem one on each side of protoxylem of centripetal xylem. Outside the centrifugal xylem is cambium and then phloem towards periphery.
(ii) Leaflet
Epidermis : Epidermis is single layered with thick cuticle. The upper epidermis is complete whereas the lower epidermis is interrupted by several stomata present only in the region of blade (hypostomatic). Upper and lower epidermis are covered by layer of thick cuticle.
Hypodermis : Just below the upper epidermis, there are several layers of sclerenchymatous hypodermis while above the lower epidermis it is present only in the midrib portion.
Mesophyll : Mesophyll is differentiated into palisade parenchyma on upper side and spongy parenchyma on lower side. Palisade tissue is made up of vertically elongated cells without intercellular spaces. Both tissues contain chloroplasts.
Vascular bundle : In the midrib there is a large vascular bundle. The vascular bundle is collateral and closed. The xylem is mesarch, i.e., diploxylic condition with centripetal and centrifugal xylem.
Transfusion tissue : On each side of the midrib in between the palisade and spongy tissues is present transfusion tissue made up of horizontally arranged tracheids which supply water and mineral to mesophyll tissue upto margins.
Reproduction
Cycas plants are dioecious and reproduce by following methods :
(1) Vegetative propagation : It occurs by means of bulbils (resting adventitious buds) which are produced on the stem in the axil of scale leaves. They break up from the parent plant and germinate to give rise to new plant.
(2) Sexual reproduction : Plant of Cycas is sporophyte (2n) and dioecious. The sexual reproduction is of oogamous type, i.e., takes place by the fusion of distinct male and female gametes. The male and female gametes are formed by the germination of micro and megaspores which are born on microsporophylls and megasporophylls. the microsporophylls are grouped together to form a compact conical structure called male cone, whereas the megasporophylls are not aggregated to form a cone, they are produced at the apex of the stem in succession with the leaves.
Male cone : The male cones are borne every year singly at the apex of the male plant. The growth of the male plant is, therefore, checked.
Later on, a lateral bud develops which pushes the male cone to one side and occupies a terminal position. The process is repeated during the formation of subsequent male cones. As such, the growth pattern of male plant is sympodial.
The male cone is a shortly stalked, oval or elliptical structure measuring about 40 – 60 cm in length. It may sometimes attain a length of 75 cm. in C. circinalis. Each cone consists of a central axis bearing numerous microsporophylls arranged in spiral manner.
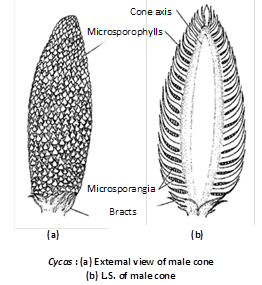
Microsporophylls : They are wedge - shaped structures with a slightly broad base. They are soft and fleshy in the younger stages. At maturity, they are hard and woody. They measure about 3 – 4 cm. in length and 1.2 – 2.3 cm. in width. They bear sori of sporangia on the abaxial (lower) surface. The terminal sterile portion of the sporophyll is called apophysis. In the apophyseal region the sporophyll gradually tapers and points upward.
Microsporangium : The microsporangia are borne in sori on the abaxial surface of the sporophyll. Each sorus contains 2 – 6 microsporangia. The number of microsporangia may be upto 700 in C.circinalis, 1000 in C. revoluta and 1150 in C.media. In between the sporangia are present uni or bicelled epidermal hair. The microsporangia are short-stalked, oval or elliptlical structures. The development of the sporangium is of eusporangiate type. Each sporangium consists of a 5 – 6 layered wall. The outer most wall layer is called as exothecium whereas the innermost layer is the tapetum.
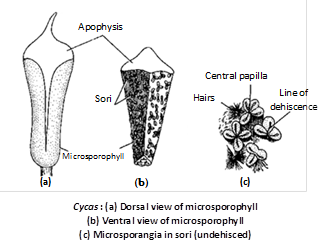
The tapetum encloses the sporogenous tissue. The sporogenous cells divide and re-divide to form the microspore mother cells or pollen mother cells (PMC). The PMC undergo meiosis to form tetrahedral tetrads of spores. The cells of exothecium develop a thickening along their radial and inner tangential walls. The cells of tapetum and inner wall layers degenerate at maturity to provide nutrition to the developing pollen grains. The wall of a mature sporangium, thus comprises exothecium only.
Megasporophyll : The megasporophylls are spirally borne in acropetal order on the female plant. Since they are loosely arranged, there is no female cone formation. Each megasporophyll is regarded as a modified foliage leaf and is about 5 – 10 inches long. In the female plant therefore, the apical meristem remains unaffected. Hence, the growth pattern in the female plant is monopodial.
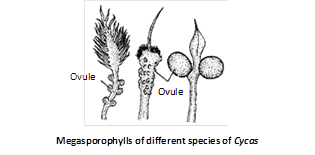
The megasporophylls is are flat, dorsiventral structures distinguishable into a proximal stalk or rachis part and a distal lamina. The margin of lamina is serrate or dentate in C. circinalis, C. beddomei and C. rumphii. In the upper part of the rachis are present 1 – 6 pairs of ovules, laterally. This number is variable in different species e.g., 1 – 6 pairs in C. revoluta, C. circinalis and only one pair in C. normanbyana.
Megasporangium (Ovule) : The ovules of Cycas are largest in nature, can be seen by naked eyes. In C. circinalis, the ovules are largest in size, i.e., about 6 cm in length and 4 cm in diameter.
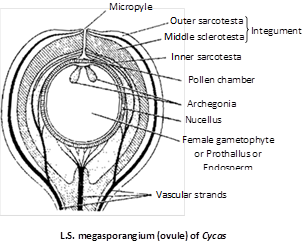
The ovules are orthotropous and unitegmic. The main body of the ovule is nucellus, covered by a single thick integument except at the top where a small opening is left called micropyle.
The integument is distinguishable into three layers, an outer fleshy layer (sarcotesta), middle stony layer (sclerotesta) and inner fleshy layer (sarcotesta). The outer and inner fleshy layers are vascularised as also the nucellus by separate bundles. The apex of the nucellus develops a beak-like process, the nucellar beak, which projects into the micropyle.
Somewhere in the deep layers of nucellus a megaspore mother cell in differentiated. It has a prominent nucleus and dense cytoplasm. It undergoes meiosis to form a linear tetrad of megaspores. Of these, three micropyler megaspores degenerate and the lowest functions. The functional megaspore has a thick papillate outer wall called exospore and a thin, fibriller inner wall, the endospore.
The gametophyte : As a result of sporogenesis, the micro and megaspores are formed. They are the first gametophytic cells. The microspores give rise to the male gametophyte whereas the megaspores form the female gametophyte. The gametophytes reproduce sexually.
Male gametophyte : The unicelled microspore undergoes two divisions of microgametogenesis and as a result three cells are formed. These three cells are serially designated as tube cell, generative cell and prothallial cell. At this stage the pollen grain is double layered. The outer wall exine is much thicker than intine. The microsporangium dehisces by a longitudinal slit and pollen grains are dispersed at 3-celled stage.
Female gametophyte : The nucleus of the functional megaspore divides freely to form a free-nuclear gametophyte. A vacuole appears in the centre. Wall formation now begins from periphery and gradually proceeds towards the centre. As a result, cellular female prothallus or megagametophyte or endosperm is formed. The 'endosperm' in Cycas is a haploid gametophytic tissue formed before fertilization. This is nutritive in function. Simultaneously, a tiny space develops on the upperside of the ovule between nuclellus and the female gametophyte due to degeneration of certain nucellar cells. This is called as archegonial chamber.
Archegonium : The archegonia are formed from the gametophytic cells lining the archegonial chamber. The number of archegonia formed in a gametophyte is variable e.g., 2 – 8 in C. revoluta, 3 – 6 in C.rumphii and 3 – 8 in C. circinalis. An archegonium consists of a two celled neck but there is no neck canal cell. There is no venter either. The egg and the ventral canal nucleus remain surrounded by the cells of prothallus. Cycas produces largest egg in the plant kingdom measuring 0.5 mm. in diameter.
Pollination : The pollination is anemophilous. The pollen grains of Cycas are light in weight and easily blown away by wind at 3-celled stage (prothallial cell, generative cell, tube cell). At the time of pollination, a large pollination drop comes out of micropylar end of ovule by disorganisation of nucellar beak. The pollen grains are entangled on this drop and as it dries, the pollens are drawn into the pollination chamber.
Post pollination changes in the male gametophyte : After a definite period of rest, the pollen grain germinates. The generative cell divides into a lower stalk cell and upper body cell. Body cell enlarges and forms several blepharoplasts, which later forms cilia.
The tube cell elongates, pierces the exine and forms a pollen tube. The pollen tube is slightly swollen and branched at tip. The pollen tube acts as haustorium absorbing food from nucellus. Body cell divides into two daughter cells and each daughter cell metamorphosis into one antherozoid or sperm or male gamete.
The male gametes of Cycas are largest (300m) in nature, visible to naked eye and are oval in form, broad (top-shaped) and naked at posterior end and spirally coiled in the anterior half with thousands of small cilia. The sperms pass into pollen tube and reach the tip of the tube.
Fertilization
After reaching the archegonial chamber, the tip of the pollen tube ruptures releasing the two male gametes. Besides, the tube also discharges a fluid having high concentration. When an antherozoid touches the neck cells, it is sucked in violently. By the time the ventral canal nucleus has already degenerated. As a result of syngamy, the zygote is formed.
The fertilization in Cycas is, therefore, siphonogamous (by pollen tube) accompanied by zoidogamy (by flagellate gametes). Thus the fertilization brings to an end of the gametophytic generation and the zygote is the initial stage of sporophytic generation.
Embryogeny
The zygote, which is the first sporophytic cell, undergoes free-nuclear divisions. A vacuole develops in the centre pushing the nuclei to the peripheral position. In the upper region there are only a few nuclei but the lower region contains numerous nuclei. This is followed by wall formation that begins from periphery and proceeds to centre (centripetal). The cellular proembryo consists of –
Upper : Haustorial region, Middle : Suspensor region and Lower : Embryonal region.
Embryo consists of radicle, plumule, 2 cotyledons, haustorium and suspensor. In cycas polyembryony is often seen as egg of almost all the archegonia are fertilized and produce embryo.
Seed formation
The mature seed of Cycas is an orange-red or reddish-brown structure. The seed is covered by a thick testa. It is sweet in taste and emits pleasant odour.
These two characteristics are responsible for the their zoochorus, (ornithochorous) dispersal. Major parts of nucellus and inner sarcotesta are used up by the developing embryo reducing them to thin, papery layers.
The embryo is distinguishable into a haustorial tip, a long suspensor, radicle, hypocotyl, plumule and two cotyledons.
Seed germination
There is hypogeal germination of Cycas seed. In germination, the radicle forms a tap root. The cotyledons remain in the endosperm under the surface of soil. The plumule grows up and forms some scale leaves and later foliage leaves. Cycas seed represents 3 generations :
Old sporophytic generation (represented by seed coat and nucellus), Female gametophytic generation (represented by endosperm), and Future sporophytic generation (represented by embryo).
Life history of Cycas is diplohaplontic. It shows heteromorphic or heterologous type of alternation of generations.
Economic importance
(1) A starch called sago is obtained from the pith of Cycas, that is why Cycas is called sago palm. In Japan starch extracted from stem of C. revoluta is used for preparing saboodana.
(2) Seed of some Cycads are used as fodder for animals.
(3) Leaves are used for making mats and baskets.
(4) Cycas is an ornamental plants.
(5) Boiled young leaves are eaten as vegetables.
(6) Extract of young Cycas leaves are used in the treatment of many skin diseases.
The decoction of seeds is used as purgative. Tincture prepared from its seeds is used by Indians in headache, nausea, bad throat, etc..



You need to login to perform this action.
You will be redirected in
3 sec

